
第580号 2006(H18).11発行
PDF版はこちら
§微量元素よもやま話[2]
モリブデン
京都大学名誉教授
高橋 英一
§被覆肥料を用いた施肥量削減茶園における収量・荒茶成分の推移
静岡県茶業試験場富士分場
小杉 由紀夫
§Vポーラスを利用した茶園での寒肥樹冠上施用の効果
チッソ旭肥料(株) 富士営業所
技術顧問 岩橋 光育
微量元素よもやま話[2]
モリブデン
京都大学名誉教授
高橋 英一
モリブデンという元素
鉛の重要な鉱石である方鉛鉱(PbS)は古代から知られており,molybdaena(ラテン名)と呼ばれていましたが,その後見出された輝水鉛鉱(MoS2)や黒鉛(=石墨C)も光沢のある灰黒色の軟らかい鉱石で外観が似ているところから方鉛鉱と混同され,長らくの間同じ名前でよばれていました。
1778年スウェーデンのシェーレ(Scheele)は輝水鉛鉱の分析を行うためこれを強い硝酸で分解したところ,硫酸と白い物質が生じました。しかし硝酸は黒鉛には作用しなかったので,この二つは別物であることが分かりました。シェーレは生成した白色物質を分離し「モリブデン土(terra molybdaena)」と名付けました。彼はまたこれが酸性物質であることから「モリブデンの酸」とも呼びましたが,同じスウェーデンの化学者べリマンはこれは新金属の酸化物に違いないと示唆しました。新金属を酸化物から取り出すには還元することが必要ですが,シェーレは鉱石を還元できる炉をもたなかったので,それを友人のイエルム(Peter Jacb Hjelm)に依頼しました。
1781年,イエルムはシェーレの「モリブデンの酸」の粉末(MoO3と思われる)を亜麻仁油で、練ってペースト状にし,密閉ルツボに入れて強熱すると,油が炭化して生じた炭素がモリブデン酸を還元し,金属を単離することに成功しました。イエルムはこの新元素を原鉱石の呼び名であったmolybdaenaに因み,molybdenumモリブデンと命名しました。その結果かつて鉛を意味したギリシャ語のmolybdosが,化学的に全く関係のない金属の名前に転化し,もとの意味は失われてしまいました。
モリブデンのクラーク数は0.0013%(37位)でホウ素より若干高いですが,土壌中の濃度は2(0.2-5)ppmでホウ素より低くなっています。モリブデンは重金属元素ですが,イオンとしては酸素と結合したモリブデン酸アニオンとして存在し,鉄やマンガンなどの重金属がカチオンの形をとるのと異なっています。土壌中では酸化鉄ゲルの表面に強く吸着される性質があるので,酸性土壌中では有効性が低く欠乏症が出やすい傾向があります。
モリブデンと植物
モリブデンと生物との関係が知られたのは,1930年にBortelsがアゾトバクターの生育にモリブデンが必須であると指摘したことに始まります。その後根粒菌,クロストリジウム,シアノバクテリアも窒素固定に依存するときモリブデンを要求することが明らかにされました。
高等植物におけるモリブデンの必須性は,1939年にArnonとStautによって偶然に発見されました。彼らは銅と亜鉛を除くのに特別注意してトマトを水耕栽培したとき,それまでの記録にない異常が現れました。それはモリブデンを含む13の元素の混合物を与えると速やかに回復しましたが,とくにモリブデンは0.01ppmで生育を正常に戻しました。
モリブデンの必須性が明らかになる前に, 二つの重要な野外で起こる病気が知られていました。一つは1908年に報告されたフロリダのカンキツの葉の黄斑病,今一つは1924年に報告されたニューヨーク州ロングアイランドのブロッコリーとカリフラワーの鞭状葉症です。これらは後にいずれもモリブデン欠乏症であることが,葉面散布試験の結果明らかにされました。
モリブデンの必須性はモリブデンを構成金属とする酵素(モリブデン酵素)である,ニトロゲナーゼと硝酸還元酵素の作用によっています。ニトロゲナーゼは分子状窒素(窒素ガス)を,硝酸還元酵素は硝酸を,それぞれアンモニアに還元する働きをし,窒素の同化に重要な役割を果たしています。
マメ科植物や非マメ科植物の根粒には,茎葉の10倍以上ものモリブデンが含まれており,モリブデンが不足すると,植物が共生的窒素固定に大きく依存している場合には窒素欠乏におちいります。またモリブデンの効果は植物が硝酸態窒素を給源にしているとき大きく,モリブデン不足の場合は体内の硝酸態窒素の含量が高まり,モリブデン施用によって減少し,生育の促進がみられます。
モリブデンの役割がこの二つの酵素の作用を通じてのみ発揮されるとすれば,植物が生育に必要なだけのアンモニアを供給されている場合,モリブデンは不用ということになります。実際施用窒素がアンモニア態の場合,モリブデン欠乏症は軽微あるいは発現しにくいことが知られています。
モリブデン欠乏症の一つに,カリフラワーやブロッコリーに発現する鞭状葉症があります。これは写真A,Bにみられるように,若い葉は主脈に沿った不規則な形の縁を残して伸び続け,主脈のみが残った鞭のような奇形な葉になります。この異常な形態は単なる窒素(アンモニア)欠乏の結果とは思えません。またこの症状は施用窒素を尿素あるいはアンモニア化合物で与えて砂耕した場合も発現します。しかしアンモニアと炭酸ガスから純粋で無菌の炭酸アンモニアをつくり,これを与えて無菌栽培したカリフラワーは硝酸還元酵素が誘導されず,鞭状葉症も現れませんでした。


この実験結果から次のようなことが推測されます。窒素源をアンモニア態に限定して与えても,自然条件=非滅菌条件のもとでは硝酸化成菌によってアンモニアから硝酸が生じます。すると硝酸還元系が誘導されるが,モリブデンがないために正常でない硝酸還元アポ酵素(金属を含まないタンパク部分)を生じ,このタンパク(の毒性)が鞭状葉症の原因ではないかというものです。この推測は魅力がありますが,毒性のしくみや,何故鞭状葉症がアブラナ科のものに限られているのかは(筆者の知る限りでは)不明のままです。
陸上植物のモリブデン濃度は平均1ppm前後,正常な生育に必要な濃度はその10分の1程度の0.1ppmと,微量必須元素中必要量は最小です(表1参照)。しかし農耕地でモリブデン欠乏は実際に起こります。一方必要量の100倍以上の10ppmを越えるモリブデンを含む場合でも,過剰害がみられることはあまりありません。
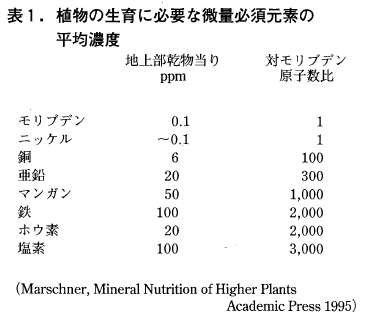
このように欠乏と過剰の濃度範囲が大きいのはモリブデンの特徴ですが,それはモリブデンが酸素酸のモリブデン酸アニオンとして行動することに関係しているとともに,後述のように草食動物に食物連鎖の上でトラブルを引き起こす原因になっています。
モリブデンと動物
モリブデンは動物の微量必須元素になっていますが,それは植物のようにニトロゲナーゼや硝酸還元酵素を介してでなく,今一つのモリブデン酵素であるキサンチンオキシダーゼ(尿酸生成に関与)によるものです。またモリブデンを除いた飼料を用いた試験で,モリブデンの必要なことは認められているが,実際に自然界で家畜のモリブデン欠乏が起こるかは明らかでなく,問題になるのはモリブデン過剰の害であり,この点でも植物と異なっています。
家畜のモリブデン中毒は世界各地でみられます。たとえばイギリス西南部のSomerset地方には,昔から”teart soil”(teart=tartは酸っぱいの意)と呼ばれる地域があり,その辺りに放牧されたウシは数日で重症の下痢(scouring)にかかり,脱毛や皮膚の脱色がおこります。その原因は長らく不明でしたが,1938年になってこれが牧草に含まれている過剰のモリブデンによって引き起こされることが明らかになりました。
この”teart soil”は中性またはアルカリ性で可溶性のモリブデンが多く,そこに生えている牧草のモリブデン含量は乾物当たり20~100ppmもあったのに対し,近くの正常な地域の牧草には3~5ppmしか含まれていませんでした。これがモリブデン中毒であることは,健康なウシにモリブデン酸ナトリウムを投与すると同様な症状が発現すること,また”scouring”にかかったウシをこのような地域から連れだし,モリブデンを低濃度しか含まない正常な牧草を与えると,治癒することから確認されました。
家畜の中でウシはもっともモリブデン耐性が弱く,ヒツジがこれにつぎ,ウマやブタは耐性が強いことが知られていますが,この耐性の違いはモリブデンの排池速度の違いによるといわれています。ウシのモリブデン中毒はイギリス以外にもアメリカ,ニュージーランド,オーストラリアなど世界各地で知られており,わが国でも島根県のモリブデン鉱山下流域に食草中の過剰のモリブデンによるウシの病変(被毛の白色化)がありました。ウシは牧草のモリブデン濃度が10ppmを越えると中毒の危険性がありますが,このモリブデン毒性は銅不足を誘発するためであるといわれ,実際硫酸銅の投与によって毒性が軽減されます*1。モリブデンと銅の相互作用については,詳しいことはまだよく分かっていないようです。
モリブデンは土壌中でモリブデン酸アニオンとして存在するので,土壌反応と溶解度の関係はカチオン性重金属の場合とは逆になります。作物がモリブデン欠乏になるのは酸性土壌においてであり(リン酸と同じように活性の鉄やアルミニウムに固定されます),ウシがモリブデン中毒になるのは土壌がアルカリ性の場合です。前述の”teart soil”の改良に硫黄華が施用されることのあるのは,硫黄が土壌中でバクテリアによって酸化されて硫酸を生じ,pHを下げる働きをするのを利用したものです。
生物進化とモリブデン
「生命とはタンパク質の存在様式である」とはエンゲルスの名言です。タンパク質は約20種のアミノ酸を原料につくられているので生物はこのアミノ酸を手に入れる必要がありますが,生物は進化の過程でその工夫をいろいろしてきました。
生命誕生以前の原始の海ができた40億年ほど前,地球には遊離の酸素はまだなく,原始大気中の窒素ガス,水素ガス,メタン,炭酸ガスなどに太陽から降り注ぐ紫外線や放電のエネルギーが作用していろいろな化合物をつくってゆきました。その中にはホルムアルデヒドやシアン化水素のような活性に富んだ物質があり,それから糖,脂肪酸,アミノ酸,核酸塩基などの低分子の有機化合物ができました*2。
これらは海水に溶け込み,紫外線による分解から守られて次第に蓄積してゆきました。そして糖と脂肪酸はそれぞれ多糖類と脂質を,アミノ酸はタンパク質をつくりました。またタンパク質のあるものは海水中の鉄や亜鉛などの重金属を取り込んで,化学反応を促進する働きのある酵素をつくりました。さらに海水中のリン酸と結合した糖と核酸塩基との複合体の中から,化学エネルギーの運搬体となるATPや,タンパク合成を指令する核酸が生じました。そしてこれらの生命物質が脂質の膜に包まれた最初の生物細胞が,三十数億年前海中に誕生しました。
つまり生命を形成する基本物質は,生物誕生に先立つこの時期(化学進化の時代)に,アミノ酸をはじめとしてすべて準備されていたわけですが,その生産効率は非常に低いものでした。初期の生物はいろいろな化学反応を促進する酵素をもつようになり,無機化合物のアンモニアからアミノ酸を自らつくるようになりました。さらにニトロゲナーゼという酵素で窒素ガスに水素を添加してアンモニアに変え,効率よくアミノ酸をつくるものも現れました。それらは窒素固定細菌として現在も活躍していますが,窒素ガスをアンモニアに変える能力は動物や植物に進化した段階で失われてしまいました。
植物や菌類は窒素固定能力はないものの,アンモニアをアミノ酸に有機化する能力は持ち続けましたが,動物はこれも失ってしまいました。その結果,自分の体のタンパク質をつくるのに,植物や菌類は窒素固定細菌に依存し,動物は植物や菌類に依存するという階層的関係が生まれました。ところで炭酸ガスから糖をつくる光合成反応は還元反応で,これには多量の水素が必要です。初期の光合成生物のうち光合成細菌は,この水素源として原始の地球環境には豊富にあった水素ガスや硫化水素を使いますが,藍藻(シアノバクテリア)は水を光分解して水素を得ています。
水は地球上に豊富に存在するので,水素ガスや硫化水素にくらべて水素源として好適ですが,問題は水の分解の結果,酸素ガスが生じることでした。酸素ガスは有機物を酸化する力が強いので,蓄積すると生体にとって危険です。しかしこれは生物を進化させる大きな推進力になりました。そしてこの藍藻の水分解型の光合成は,その後植物にうけつがれて発展して行きました。
さて酸素ガスの発生によって環境が酸化的になってから,窒素ガス→アンモニア→アミノ酸→タンパク質というメインルートにバイパスができました。それはアンモニアを酸素ガスで酸化して硝酸にかえる硝化細菌が出現したためです。硝化細菌は酸化の途中で遊離するエネルギーを利用しています。このバクテリアの働きでアンモニアがどんどん硝酸になってしまうと,植物はアンモニア欠乏すなわち窒素欠乏になってしまいます。それでは困るので植物は硝酸を再びアンモニアにかえて利用するシステムを開発しました。そのとき鍵酵素としての役割を演じているのが硝酸還元酵素です。
モリブデンはこれらの酵素(ニトロゲナーゼと硝酸還元酵素)の金属成分として,生物進化の中で二度にわたって重要な役割を演じました。最初は窒素固定細菌の出現した生物進化の初期の時代であり,二度目は環境中の酸素濃度が増加し,好気呼吸を行う生物が現れてからあとの時代です。アンモニアが硝酸に酸化されるようになったのは,酸素濃度が現在の百分のーを超えてからのことであるはずなので,モリブデンが硝酸還元の役割をもつようになったのは今から15億年ほど前,最初に窒素固定に関与してから20億年近くたってからということになります。そしてその役割は2回ともタンパク合成の出発物質であるアンモニアを供給するところにあったことは興味深いものがあります。
*1 硫酸銅の硫酸根がモリブデン排出を促進し,銅/モリブデン比を高める効果もあるといわれる。
*2 1953年シカゴ大学で新分野を切り開く実験が行われた。ノーベル賞受賞者ハロルド・ユーレイの下で博士課程の学生であったスタンレー・ミラーは,原始大気をまねてアンモニア,水蒸気,水素,メタンを混合し,その中で一週間ほど放電を行った。するとアミノ酸であるアラニンとグリシンその他いろいろな有機物質ができた。・・・・その後生体の複雑な分子を構成する化合物のほとんどは,ガスやミネラルの溶液にスパークや紫外線照射などいろいろな方法でエネルギーを与えることによって合成された。
マルグリス・セーガン共著,田宮信雄訳
ミクロコスモス東京化学同人1995
被覆肥料を用いた施肥量削減茶園における
収量・荒茶成分の推移
静岡県茶業試験場富士分場
小杉 由紀夫
1.試験の背景・ねらい
茶栽培においては窒素肥料が環境ヘ負荷を与えていることから,施肥量を削減した施肥技術が求められて来ました。一方,長期に渡り施肥量削減した場合,収量や品質がどのように変化するのか明らかにすることも必要でした。そのため,静岡県茶業試験場では,現地農家の協力のもと,施肥量を削減した場合の収量や品質に対する影響について県の中部,西部地域で継続的な調査を行ってきました。しかし,本県の中,西部地域の多くの茶園が赤黄色土壌であり,また茶業試験場のある牧之原台地も同じ土壌であることから,試験データは赤黄色土壌条件下のものでした。
一方,静岡県の富士川以東の茶園は富士山の火山灰に由来する黒ボク土壌であり,赤黄色土壌での施肥量削減の結果がそのまま活用できるのか判然としませんでした。そこで,黒ボク土壌地帯にある当分場では,平成10年から試験開始前平成9年の標準的窒素施用量75kg/10aを基準に,54kgと31kgに施用量を削減した場合の生葉収量や荒茶成分への影響を継続的に調査しました。
2.試験内容
富士分場の茶園(品種’やぶきた’,19年生)を用いて,1区20㎡,2反復で平成10年2月の春肥から処理を開始しました。各区の年間の施肥体系は表1のとおりです(以下,それぞれの区のNを省略する)。なお,31kg区では春と秋に施用する配合肥料に被覆肥料(エコロング複合S301-70)を重量比で14%加えてあり,年間窒素量の約1/3は被覆肥料由来としました。調査は一番茶,二番茶の生葉収量と枠摘み(30×30cm)による収量構成要素,少量製茶機(1kg機)により製造した荒茶の成分分析及び官能検査,土壌の理化学性調等を行いました。
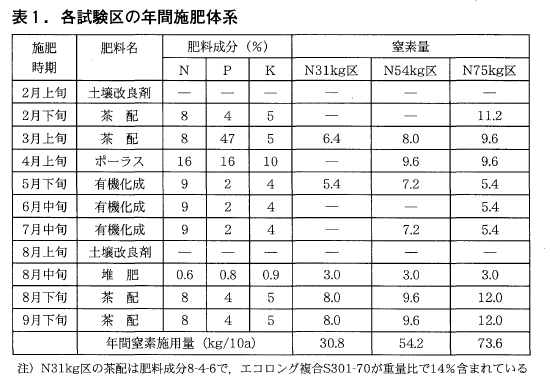
3.調査結果
(1)生葉収量
試験開始3年目となる平成12年からの一番茶及び二番茶の各区収量の平均値の推移を表2に示しました。一番茶では,年により収量に差は見られますが,統計的には区間に差は見られず,いずれの年も31kg区,54kg区とも対照の75kg区と変わらない結果でした。31kg区では窒素削減が大きいにもかかわらず,平成17年を除けばいずれの年も75kg区を上回るような収量でしたが,この要因の1つは被覆肥料を用いたためとも考えられます。
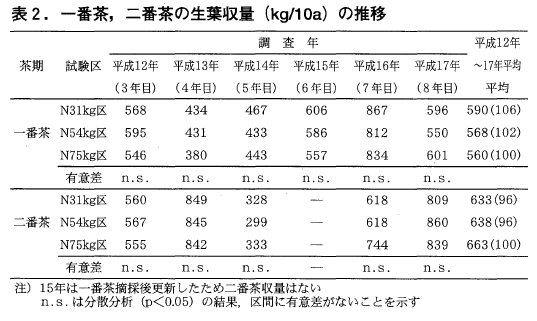
また,二番茶も一番茶同様3つの区の間に統計的な差はありませんでした。ただ,一番茶では31kg区,54kg区とも6年間の平均値が75kg区をやや上回るような値でしたが,二番茶では5年間の平均値が75kg区をやや下回るような値となってしまいました。この要因としては中切り更新した翌年(平成16年)の収量が75kg区に比べ少なかったことが影響しているように思われました。
(2)枠摘み調査
一番茶の枠摘み調査結果を表3に示しましたが,新芽数(芽の数),百芽重(芽当たりの重量),出開度(芽の生育程度)とも収量同様,区間に統計的な差はありませんでした。また31kg区はいずれの調査項目も,6年間の平均値は75kg区をやや上回る数値となっています。
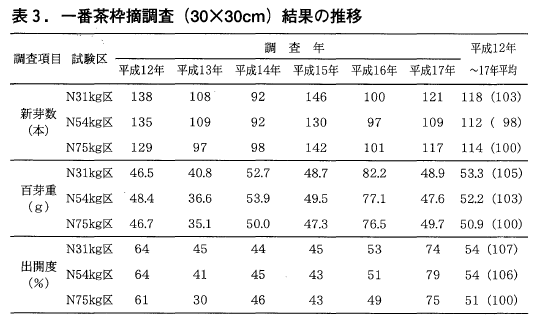
一方,二番茶の枠摘み調査結果も,一番茶同様いずれの年も区間に統計的な差は見られませんでしたが,新芽数,百芽重の6年間の平均値は,31kg区,54kg区とも75kg区に比べ低い値となっております(表4)。
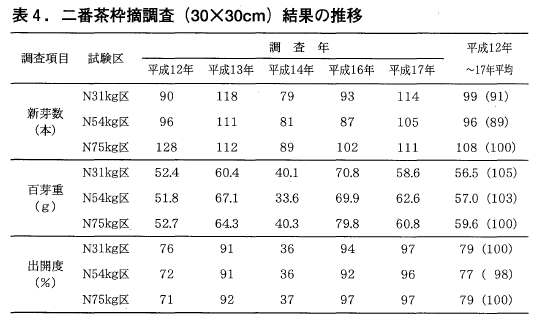
新芽数は平成12年と中切り更新した翌年(平成16年)の減少が,百芽重は平成16年の減少が影響を与えているように考えられます。
(3)荒茶成分
近赤外分光分析法による一番茶荒茶の全窒素,遊離アミノ酸,粗繊維の含有率はいずれの年も,区間に統計的な差は認められませんでした。しかし,全窒素,遊離アミノ酸では平成15年から31kg区,54kg区とも75kg区に比べわずかに低い傾向が見られます(表5)。
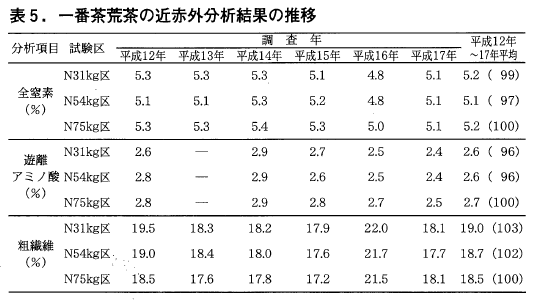
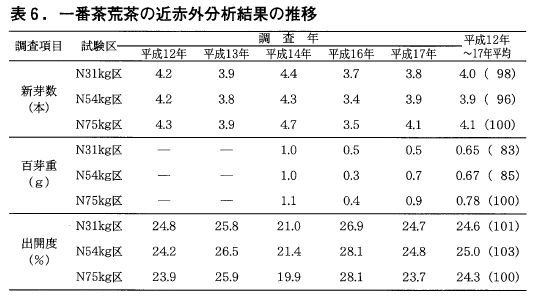
一方,粗繊維は処理開始当初はわずかに多い傾向が伺えましたが,7,8年目はほぼ同量になってきたように思われます。また二番茶でも5年間の平均値は一番茶と同様の傾向が伺えます(表6)。
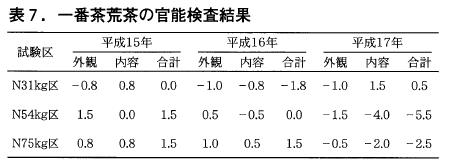
(4)官能検査
平成15~17年の一番茶荒茶の官能検査では,平成15,16年は75kg区の点数が他の2区より優れる傾向にありましたが,平成17年は逆の結果となりました(表7)。官能検査では製造によるサンプル間の誤差が出やすく,区間の差は判然としないと思われました。
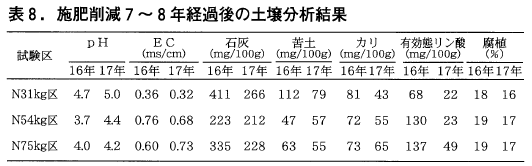
(5)土壌の理化学性
処理後7,8年経過した平成16,17年の土壌分析(表8)の結果をみてみると,31kg区では,pHは他区に比べ高く,またEC値は低くなっています。一方,54kg区は75kg区と変わりません。土壌改良剤の処理量が同じ31kg区と54kg区では石灰と苦土の値にも差が見られ,54kg区では両成分が窒素とともに流亡しているために値が小さくなっていることが推察されます。カリは年により数値が異なるものの,区間に大きな差はありませんが,有効態リン酸は施肥量が多い区の方が値が高く土壌に蓄積していることがわかります。
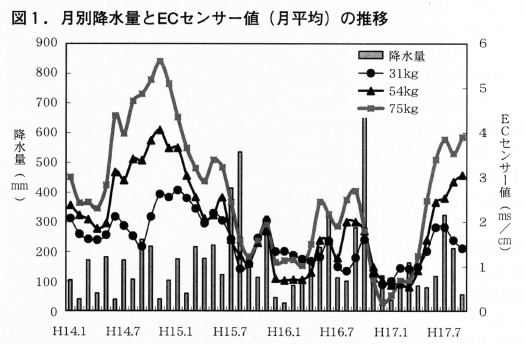
月別降水量とECセンサー値の推移を図1に示しました。試験ほ場に埋設されたECセンサー値は施肥量が多い区で数値が高い傾向がみられます。しかし,夏期に降水量の多かった年の秋~冬期は区間に差は少なく,ECセンサー値が逆転する月も見られます。このことから,施用量が多くても降水量が多いと,窒素等の塩基の流亡が多いことが推測されます。一方,31kg区や54kg区の窒素削減区では被覆肥料を配合することにより,降雨による窒素分の流亡を抑え,一定の肥効を確保しているものとも考えられます。
3.まとめ
平成12年から平成17年までの6年間の調査データを見てみると,一番茶収量は窒素の施用量を対照1/2以下の31kgに削減しても,黒ボク土壌においては変わらないことがわかりました。また枠摘み調査結果が示しているように,施肥量を削減しても収量を構成する要素である新芽数や百芽重,また出開度などは変わらないため,収量そのものも変わらないと考えられます。一方,二番茶は統計的には3つの区間に差は見られませんが,削減区では,処理後5年目位から75kg区に比べ収量がやや少ない傾向が見られ始めたように思われます。枠摘み調査の結果では,新芽数と百芽重が少なくなる傾向にあったためと考えられます。
荒茶では,一番茶,二番茶とも調査したいずれの成分も統計的には削減による区間の差は見られません。しかし,75kg区との比較では,削減区の一番茶で遊離アミノ酸の含有率が6年目以降わずかに少ない傾向が伺えました。ただ,官能検査の点数と比較してみると,必ずしも,全窒素や遊離アミノ酸含有率が高い方が官能検査の結果が高いわけではありません。これらのことから考えると,今回の試験では,施用量の削減により荒茶品質が低下したとは考えられません。
施肥量削減の大きな目的の1つは,肥料成分の流亡による環境への負荷を少なくすることです。削減区では,pHやEC値が対照区に比べ差が見られ,また土壌改良剤の処理量が少ないにもかかわらず,土壌中の石灰や苦土の量は対照区に比べ多くなっています。このことは,施肥量を削減するとともに被覆肥料を用いることにより,環境中ヘ流亡する肥料成分が少なくなり,さらに肥料成分の吸収効率を高めることを示していると思われます。
今回の調査結果は,被覆肥料を使った継続的な施肥削減による収量や品質に対する影響を現象として捉えた試験で,それなりの効果が認められましたがその要因解析には至っていません。そのため,どこでも活用できる技術とはなっていません。今後は,茶の肥料成分の吸収メカニズムや生育との関係など,既存の情報と施肥技術を組み合わせて,多くの場面で活用可能な技術の開発が必要と考えます。
Vポーラスを利用した茶園での
寒肥樹冠上施用の効果
チッソ旭肥料(株) 富士営業所
技術顧問 岩橋 光育
1.はじめに
前報1)において,茶樹への春期の早めの窒素供給を目的に速効性肥料である’あさひVポーラス(以下,ポーラスと記す)’,を寒肥,春肥Ⅰ及び芽出し肥として茶園に施用しその効果を調査した結果,①一番茶新芽の生育は,寒肥(ポーラス)区>春肥Ⅰ(ポーラス)区>芽出し肥(ポーラス)区>慣行区の順で良好であった。②ポーラス施用に伴う生葉売上収益は慣行区に比べ増加が認められた。
残された疑問点として,ポーラスでなく他の肥料,例えば硫安でも同じ効果が得られるのではないかという疑問点の解明の為,2006年同じ試験ほ場で再試験を行った。
一方,茶樹の施肥管理はうね間に限定されており,このことが吸収根城の局所限定,根の濃度障害,肥料成分の溶脱など肥料の利用効率の向上を抑制するなどの問題点を抱えている。また,茶樹の樹冠下の根群域は広く2),吸収根の活性が高く3),かつバラツキはあるが雨水による樹冠下への落下4)などにより樹冠下施用による吸収効率の向上が期待される。
そこで,ポーラスが水に溶けやすく,茶樹成葉への直接の影響が少ないという特性,さらに雨水による樹冠下への肥料成分の供給,それに伴う吸収増を期待し,寒肥としてポーラスを茶樹冠上から散布し,その施用が茶樹の一番茶新芽の生育,収量及び品質に及ぼす効果を調査したので合わせて報告する。
2.試験方法
1)ほ場試験
試験区の構成は表1に示したとおりであり,ポーラス樹冠上施用区,ポーラスうね間施用区,硫安うね間施用区及び無施用(慣行)区の4区を設けた。
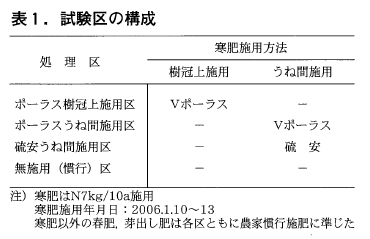
試験は掛川市大野(褐色森林土),富士市石坂(黒ボク土),川根本町藤川(レキ質土)及び菊川市倉沢(赤黄色土)の4試験ほ場で実施した。但し,菊川市倉沢の試験ほ場ではポーラス樹冠上施用区は設けなかった。
試験はいずれも‘やぶきた’成木園で2006年に行い,処理区の規模は1区14~31㎡の範囲で,富士市での試験のみ2反復で実施した。寒肥として使用した肥料は’あさひVポーラス’(窒素・りん酸・カリ:16(アンモニア態窒素13.5,硝酸態窒素2.5)-7-12)と硫安で1月初中旬に窒素として7kg/10a施用した。寒肥以外の春肥,芽出し肥料の施用は農家慣行に準じた。
調査は一番茶期に枠摘み(20×20cm,1区6ヶ所)し,新芽の生育調査を行い,また新芽は乾燥後,近赤外分光光度計による測定(全窒素,遊離アミノ酸,テアニン,粗繊維,タンニン,カフェイン)を行った。さらに,ポーラス樹冠上施用区,無施用(慣行)区の樹冠下土壌(深さ0~5cm)を3月中旬に採取し分析した。
2)カラム試験
2006年,黒ボク土の茶園から樹冠下の整せん枝有機物を採取し,洗浄処理した有機物を塩ビ製パイプ(直径10cm,長さ20cm)に充填(厚さ5,10,15cm)し,一晩放置した。翌日表層に供試肥料(ポーラス,硫安,N7kg/10a)を散布した後,上部から散水処理(150,250,350mm)を行い,カラム底部からの流出水を採取し無機態窒素,りん酸,カリについて分析し,各肥料成分の流出量を調査した。
3.結果及び考察
1)一番茶新芽生育
表2に一番茶新芽の生育調査結果を示した。新芽重は無施用(慣行)区100に対し,各試験ほ場平均でポーラス樹冠上施用区,ポーラスうね間施用区共に109と高く,次に硫安うね間施用区は104であった。
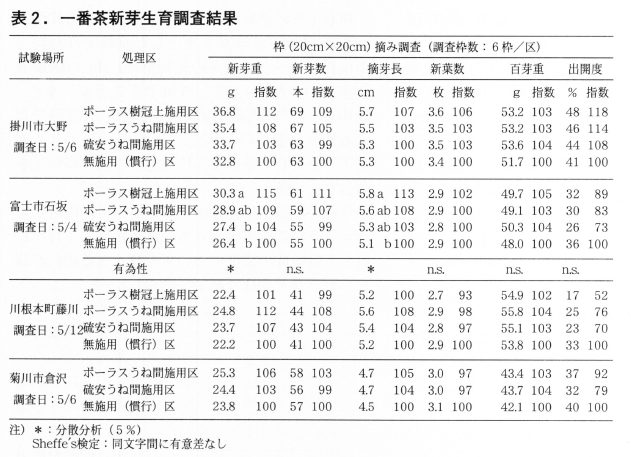
新芽数はポーラス樹冠下施用区,ポーラスうね間施用区共に106,硫安うね間施用区は100であった。摘芽長はポーラス樹冠下施用区107,ポーラスうね間施用区106,硫安うね間施用区103であった。百芽重は寒肥を施用した各施用区の平均で103と高くなる傾向が認められた。
ここでポーラス樹冠上施用区の新芽に注目すると,川根本町試験ほ場での結果は他の2試験ほ場と異なり,ポーラス樹冠上施用区で新芽の生育が抑制され施用効果が認められなかった。新芽が生育抑制された原因としては,当該試験ほ場が標高280m(他の2試験ほ場の標高は約80,90m)の山間地に位置し,寒肥施用後に最低気温-7℃以下を1月は1回,2月は3回(2006年 川根本町(標高290m)アメダスデータ)遭遇したことにより,ポーラス樹冠上施用区のみで冬芽がポーラスと接触したことによる影響と凍霜・寒さによる影響との複合作用により結果として生育抑制を受けた為と考えられるが明確ではなく,今後検討する必要がある。
2)一番茶生葉収量
図1に処理の違いによる推定生葉収量を示した。なお,推定生葉収量は新芽重,摘採面幅及びうね幅から試算した。各試験ほ場において生葉収量が多いのは,ポーラス樹冠上施用区>ポーラスうね間施用区>硫安うね間施用区>無施用(慣行)区の順であった。ポーラスの樹冠上施用区での無施用(慣行)区に比べての差は,川根本町試験ほ場では4kg/10aと小さかったが,他の2試験ほ場では各々84kg,82kgの収量増となりポーラスの樹冠上施用の効果が認められた。
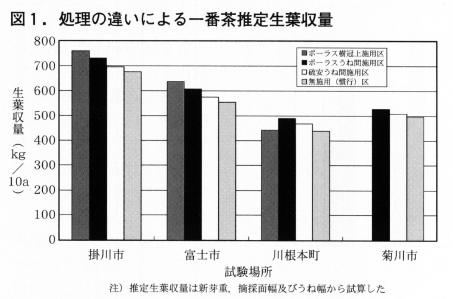
次にポーラスうね間施用区は,無施用(慣行)区に比べて平均48kg/10aの収量増であり,2005年の試験結果(平均56kg/10a増)と同様に寒肥としての施用効果が認められた。硫安うね間施用区は平均21kg/10aの収量増となり施用効果は小さかった。
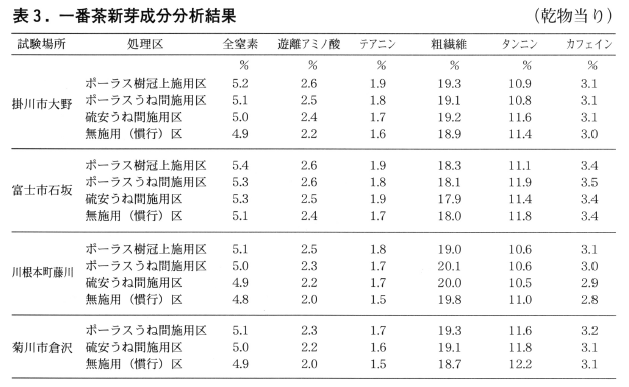
3)一番茶新芽の成分分析
表3に一番茶新芽の成分分析結果を示した。遊離アミノ酸,テアニンなどの茶の旨味成分合有率は寒肥を施用した区では無施用(慣行)区に比べ高くなる傾向が認められた。その中で特にポーラス樹冠上施用区,ポーラスうね間施用区で高くなった。
樹冠下への肥料施用の効果について,寿江島ら5)は15N標識窒素硫安を施用した試験でうね間施用に比べ一~三番新芽の乾物重が1.4倍,15N標識窒素量の分配が1.6倍と増加することを,また,鳥山ら6)は15N標識被覆尿素を施用した試験ではうね間表層施用の1.9倍となるなど,いずれも樹冠下の根からの吸収増に伴う肥料の利用率向上を報告している。
今回の試験結果でもポーラス樹冠上施用区がポーラスうね間施用区よりも一番茶新芽の生育が良好で,旨味成分濃度においても高くなる傾向が認められるなど同様な結果が得られた。
4)カラムからの肥料成分の流出
図2に充填有機物の厚さの違いによるカラムからの肥料成分の流出状況をしめした。
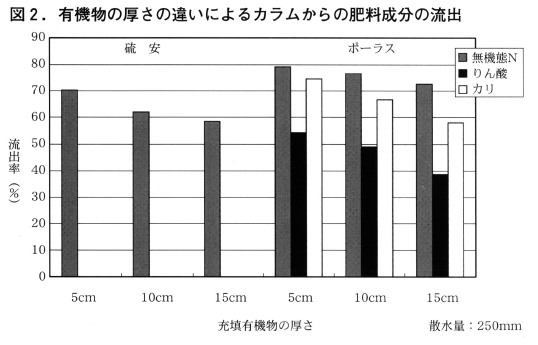
整せん枝有機物の上部に散布された硫安,ポーラスともに散水によりカラムからの流出率は有機物の層が5cm,10cm,15cmと厚くなるほど小さくなる傾向が見られ,厚さ10cmで散水量250mmでは硫安の流出率は62%,ポーラスでは無機態窒素が77%,りん酸48%,カリ67%であった。また厚さ5cmで散水量350mmでは硫安の流出率が73%,ポーラスでは無機態窒素が83%,りん酸58%,カリ76%であった。(関連図表なし)このように整せん枝有機物に移行した肥料成分は散水に伴い大半が下層へと移動,流出する傾向が認められた。
5)樹冠下土壌分析
表4にポーラス樹冠上施用後の樹冠下土壌分析結果を示した。
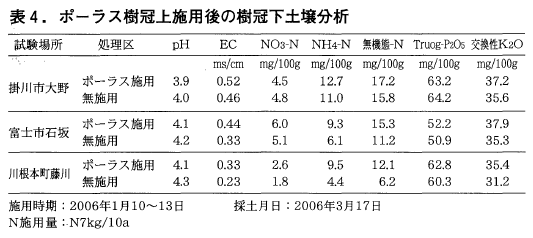
各試験ほ場ともに無施用(慣行)区に比べポーラス施用区において風乾土100g当たり無機態窒素で2~6mg,Truog-りん酸で1~3mg,交換性カリで2~3mg程度濃度の増加が認められた。1月中旬に樹冠上施用されたポーラスは,雨水による株元土壌への浸透,風や降雨による樹冠面からの樹冠下整せん枝有機物層への落下,さらにその後の降雨(富士地区降水量542mm,2006年1月~4月 アメダスデータ)により樹冠下土壌に浸透したと思われる。
さらに土壌に浸透した窒素,りん酸,カリの各肥料成分が樹冠下に分布する根により吸収され,このことがポーラス樹冠上施用区の新芽の生育,成分増加に寄与した可能性は高いと考えられる。
6)ポーラス施用に伴う経済的効果
表5にポーラス施用に伴う投資効果試算結果を示した。ポーラスの樹冠上施用区の施用効果を見ると,10a当りの生葉売上高は無施用(慣行)区に比べ3試験ほ場平均で26,595円(各々1,941,38,524,39,319円)の収入増であった。さらに肥料代,散布作業賃金を差し引いた収益では川根本町試験ほ場では無施用(慣行)区に比べ6,042円の減収となったが,他の2試験ほ場では31,335,30,540円と寒肥としてのポーラス樹冠上施用で概ね期待に近い結果が得られた。
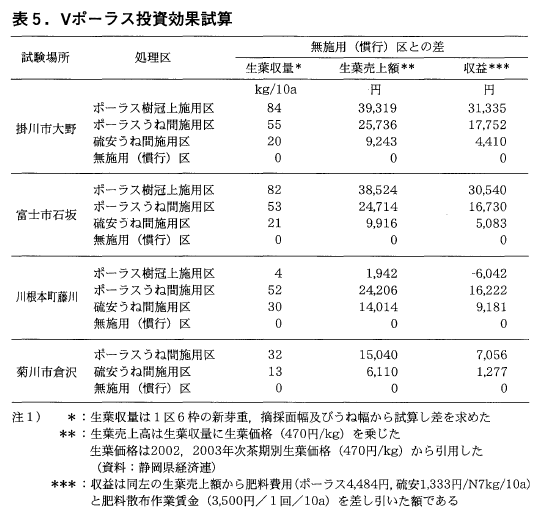
一方,ポーラスうね間施用区では無施用(慣行)区に比べ収益は4試験ほ場平均で14,440円の収入増となったが,前年(2005年)の寒肥試験の結果(平均18,321円増)に比べ収入はやや低下した。
このように前年の試験結果に続き,寒肥の施用効果が認められた。その中で最も効果が得られたのがポーラス樹冠上施用区であり,次にポーラスうね間施用区とポーラスの寒肥としての施用の効果が認められた。また,硫安うね間施用区では収益平均が4,988円の収入増となった。
これらの事からポーラスうね間施用区と硫安うね間施用区との収益性の比較では10a当たりの収益は平均で9,500円程度の差があり,寒肥の施用に当たってポーラスの硫安に対する優位性が認められた。
一般に摘採の適期は出開度70%程度(佐波7))とされているが,今回の各試験ほ場での生育調査時の出開度が17~48%と若干前倒しで調査を行ったことを考慮すると,各ほ場での現場実態では収量,収益ともに更なる増加が予想される。
7)ポーラス樹冠上施用にあたっての留意点・問題点
寒肥としてポーラスを樹冠上施用する場合,その効果を十分に発揮させる為には,適応地域の選定が必要である。今回,新芽生育抑制を受けた川根本町試験ほ場が標高280mの山間地で1,2月の最低気温が-7℃以下(最低気温平年値,1月-2.4℃,2月-2.0℃(アメダスデータ))を3回遭遇した地域であるのに対して,施用効果が認められた他の掛川市と富士市の2試験ほ場の標高は各々約80m,90mで2006年1,2月の最低気温が-4°C程度(富士地区の最低気温平年値,1月1.1℃,2月1.5℃(アメダスデータ))であった。これらのことからポーラス樹冠上施用を導入するに適した地域の条件としては,標高100m以下,もしくは最低気温平年値が1℃以上に該当する地域が適当であろうと考えられる。(ちなみに静岡県を代表する牧の原台地に位置する県立茶業試験場(菊川市)の標高は190m,最低気温平年値は1月0.6℃,2月0.6℃(アメダスデータ)である)
また,樹冠上施用後の樹冠下への肥料成分の供給を高め,導入効果を高める為には,スプリンクラーなどでかん水を行うことがより効果的と考えられる。
今後の残された問題点として,更なる樹冠下における利用率向上のために,樹冠下ヘ1年間を通じた肥料の安定供給,さらに茶樹更新期間を通じての安定供給が必要である。そこで新しい施肥体系のために,今後,緩効性肥料を中心とした樹冠上施用と従来のうね間施用を組み合わせた新たな施肥試験を予定している。
4.まとめ
前報1)での残された疑問点,寒肥としてのポーラスの施用効果がポーラスでなく硫安でも同じ効果が得られるのではないかという疑問点の解明の為,2006年同じ試験ほ場で再度の寒肥試験を,またポーラスの寒肥としての樹冠上施用が茶樹の一番茶新芽の生育,収量及び品質に及ぼす効果を県内4ほ場で調査した。
1)一番茶新芽の生育は,ポーラス樹冠上施用区>ポーラスうね間施用区>硫安うね間施用区>寒肥無施用(慣行)区の順で良好であった。
2)ポーラス施肥区の一番茶新芽の遊離アミノ酸,テアニンなどの旨味成分濃度が無施用(慣行)区に比べ高くなる傾向であった。
3)有機物充填カラムからの肥料成分の流出は250mmの散水で50%以上であった。
4)ポーラス樹冠上施用に伴う樹冠下土壌の肥料成分(窒素,りん酸,カリ)は無施用(慣行)区に比べ風乾土100g当たり数mg増加した。
5)寒肥施用に伴う生葉売上収益は無施用(慣行)区に比べ増加することが認められた。その中でポーラス樹冠上施用区では山間地試験ほ場(標高280m)を除いて高い収益(30,000円以上/10a)増が得られた。またポーラスうね間施用区と硫安うね間施用区との収益性の比較では,ポーラスの硫安に対する優位性が認められた。
6)ポーラス樹冠上施用の導入地域の条件としては,標高100m以下,もしくは最低気温平年値1℃以上に該当する地域が適当であろう。
5.引用文献
1)岩橋光育:農業と科学,1,10~14(2006)
2)青野英也 他:茶技研,56,10~29(1979)
3)小泉豊 他:昭和58年度原子炉の大学共同利用研究報告(1984)
4)辻正樹 他:茶研報,94,7~14(2002)
5)寿江島久美子 他:鹿児島県茶研報,13,12~55(1999)
6)鳥山光昭:九農研,58,81(1996)
7)佐波哲治:茶の栽培と利用加工,(株)養賢堂,142(1994)