
第741号 2022(R04).06発行
PDF版はこちら
§トマトの昼夜間における養水分吸収とその移行
元 岡山大学大学院 自然科学研究科
桝田 正治
§土のはなし−第12回
堆肥の効果の現れ方と土の条件
−土の黒さが決め手−
ジェイカムアグリ株式会社
北海道支店 技術顧問
松中 照夫
トマトの昼夜間における養水分吸収とその移行
元 岡山大学大学院 自然科学研究科
桝田 正治
1.はじめに
地球上の生物は1日を単位とした昼−夜の自然サイクルの中で生きている。この地球は1年に1回太陽の周りを巡るが,南極と北極を軸とする自転軸が,いわゆる公転面から23.5度傾いているため,日本のような中緯度地帯では昼夜の長さと温度が変化し四季を生む。植物の多くはこの日の長さや温度に反応して花を咲かせ実を結ぶ。赤道付近の低緯度地域は,自転と公転が同一軌道上にあるため四季は生じず,1日約12時間−12時間で昼夜を繰り返す。この地帯では雨季と乾季が生じ,植物の多くは雨に反応して花を咲かせる。砂漠には多肉植物が多く,サボテンの様に水を貯めるものやバオバブに見られるように幹に水を貯め乾季に葉を落とし雨季には葉を茂らせるものもある。このように地球上の植物はまさに光,温度,水を含む自然環境に適応しつつ進化してきたといえる。
地上部に見られる様々な形態も個体の維持があってこそ成り立つもので,その生存は根の存在なしにはあり得ない。根は地上部から光合成産物を受け取り,水や養分を地上部に送る。その水の流れを図1に示したが,根は土壌に引き付けられた水〈水ポテンシャルと称し,値は常にマイナスとなる〉を吸い,水ポテンシャルのより低い茎から葉へと水を上げる。上げるというよりも水は葉の水ポテンシャルによって生じる蒸散力によって引っ張り上げられるのである。水は葉から蒸発し気化熱となって葉面を冷やす。この時,イオンの形で水に溶けている様々な養分は各部位に分配されることになる。植物体内の水流は昼と夜で大きく異なるが,これに伴う吸水の量ならびに養分の量,そしてその成分バランスはどのようになっているのであろうか?
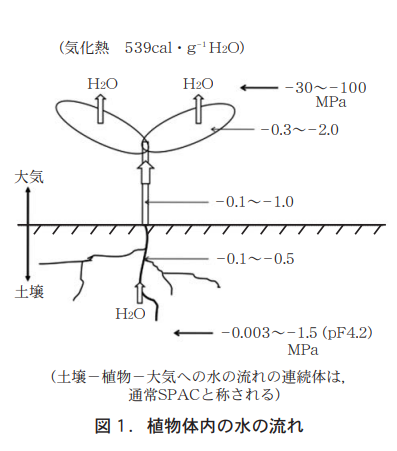
本論は地上部の昼夜環境に対応して根の養水分の吸収と移行はどう変化するのか,主に筆者がこれまで行ってきた研究を中心に文献を引用しつつ総説的に論述したものである。
2.昼夜間における茎,根の伸長と果実肥大
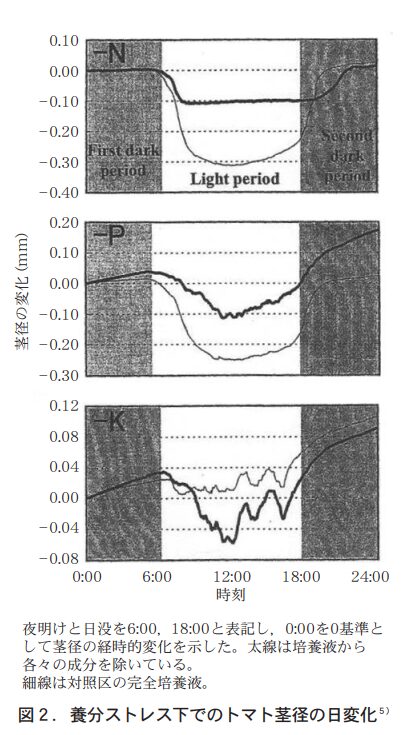
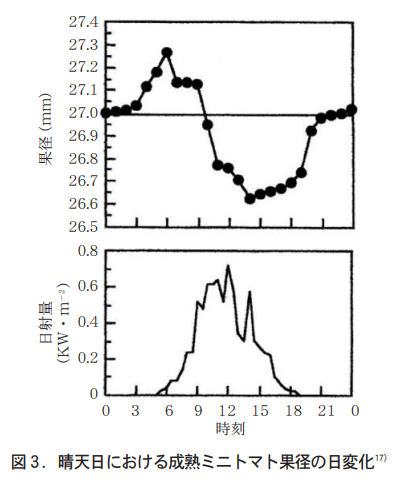
トマトの茎の伸長は,日単位で見ると昼間よりも夜間の方が大きく70〜80%は夜間に行われ25),茎の直径は昼間に縮む(図2) 。昼間に縮む現象はブドウでも端的に示されている4)。これは茎組織での水ポテンシャルが昼間に低下し細胞の膨圧が低下することによる。トマトの果径についても,昼間に減少し13時頃には最低となることが示されている(図3) 。
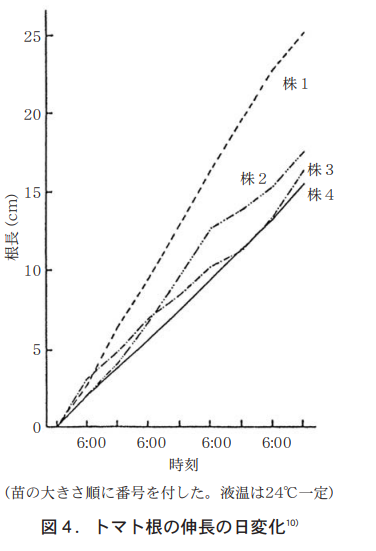
一方,根の伸長については,筆者がトタン板に傾斜をつけてトマト苗を置き,24℃の養液を連続して流し,朝6時と夕方6時に黒色シート下で伸びる根を測定したところ,根は昼夜に関係なくほぼ直線的に伸長し,若干の個体差が見られるものの時間当り約3mm前後となる(図4) 。
前報の総合考察において大学入試センター試験問題を引用したように12),根は地上部から送られてくるシグナルによって水や養分の吸収に応えている。例えば,大玉トマトが第3果房まで着果したとき果実を摘果すると,12日後には根の乾物重は対照株の3割増し,培養液のNO3−N,Ca,Mgの濃度は急落する。これは摘果によって,これら5) 成分の吸収濃度(n/w,後述)が急激に増すことを意味する。しかし,PとKの濃度に変化は見られない13)。成分によって吸収の速度が異なってくる理由については今なお分かっていない。
近年,塚本ら23)は非破壊で植物生体内の物質動態を可視化できるPositronemitting Tracer Imaging System(PETIS) を用いて,トマト側枝葉に処理した11CO2の移動画像から側枝葉の果実への寄与率を求めているが,本手法は摘葉や摘果に伴う根での養分動態解析などにも適用できるものと思われ,今後大いに期待されるところである。
3.昼夜間における葉と果実の水ポテンシャル
−裂果の様相−
先に述べたように,植物の葉から蒸散することによって体内の水ポテンシャルは低下する。それが吸水力を生み出し,水は導管の中をマスフローとして引き上げられていく。通常の水ポテンシャルは,昼間に低下し夜間には上昇する。しかし,夜間でも水ポテンシャルは常に負で正になることはない。図5に示したように,トマトの葉の水ポテンシャルは潅水量にかかわらず14時頃に−1.0MPaで最低となり真夜中に最高となる。果実の裂果は,とくに乾燥後の雨で多発することが広く知られており,対策として雨よけ栽培などの技術が普及している。
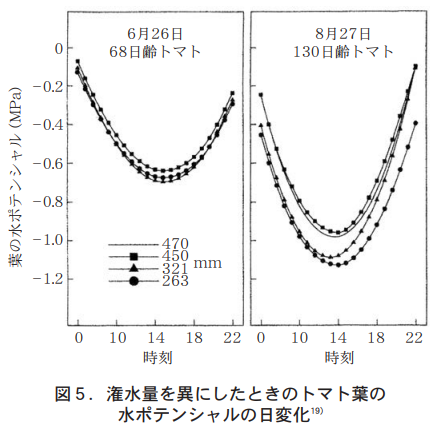
太田17)は,ミニトマトの裂果について調べ,早朝の5〜7時に多く発生すること,この頃に果実の横径が増すことから,裂果には圧ポテンシャルの増大による果実の膨張が関係しているとした。さらに同氏は夜間に葉での蒸散を促進すれば果実への水流入が少なくなるとの考えから,夜間に6000luxの照明を行ったところ,裂果は対照区の40%以下に抑えられたとしている。実験条件としての土耕と水耕,また普通トマトとミニトマトなどの違いを考慮したとしても,図5に示したように,葉の水ポテンシャルが最高値となる真夜中の時間帯と裂果が発生する朝方時間帯を考えるとき,両者には約6時間のタイムラグが生じていることが分かる。この6時間における果肉,果皮組織への水流入の動態が重水トレーサーなどにより明らかになれば,裂果のさらなる制御技術も生まれるであろうと期待される。
4.昼夜間における養分の吸収濃度(n/w)と養分移行
筆者らは,園試標準培養液においてトマトの昼夜における養分吸収濃度(n/w)を求めたところ,昼はどの成分も培養液の濃度に近い値となるが,夜は昼間の2.2倍から3.7倍となることを1984年園芸学会春季大会において発表した。同大会において,寺林らも同様な結果を報告している。ここでいう吸収濃度とは,1970年代に山崎ら24)によって提唱された概念で,広く一般に使用され野菜の培養液管理の指針とされてきた。これは,作物の吸収した成分量(n)を水量(w)で除した値で,通常me/ℓで表記される。春季に昼用〈6:00−18:00〉と夜用〈18:00−6:00〉のポットに標準培養液を入れ,同一株を移し替えること5日間,この養水分のn/w値を図6に示した。昼間のn/wは当初の培養液濃度に近い値となるが,夜間のn/wはどの成分も培養液濃度の2〜3倍で,特にPは6倍近くになることが分かる。寺林ら22)も,夏季に昼間7:00−19:00,夜間19:00−7:00として4日間株を移し替え調査した結果,Pの夜間吸収率は他の成分に比べて非常に高くなることを認めている。
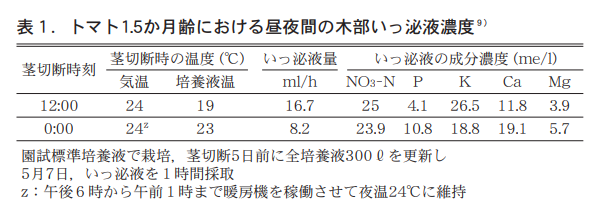
ここで当初の培養液Xℓ,終了時の液量Yℓ,この間の水面蒸発量Zℓ(ブランク区) ,処理前の濃度a,処理後の濃度bと仮定すると,n/wは(aX−bY) / (X−Y−Z)で示される。このn/wが夜間に高くなるということは,地上部へ移行する木部液の濃度も高くなることを強く示唆する。そこで,夜間の気温を昼間12:00の気温と同じに設定して,1.5か月齢トマトの茎を地際から10cmの位置で切断し,木部いっ泌液を採取分析したところ,夜のいっ泌液量は昼の半量であり,NO3−NとKの濃度は昼に高く,P,Ca,Mgは夜に高いことが分かった。特にKは昼に夜の1.4倍,Pは夜に昼の2.5倍高くなるのが特徴的であった(表1) 。
さらに,同一培養液でトマトを育て24時間にわたり1時間毎に一株ずつ茎を切除して15分間採取したいっ泌液を分析した結果,成分濃度の回帰曲線においてKは朝方から正午にかけて,Pは夕方から午後10時にかけて上昇することが明らかとなった14)。
茎を切除することによって木部液の流速はどの程度に落ちるのか? 茎切除予定部位の数cm下位にヒートパルス式サーミスターを差し込み,2か月齢のトマトの木部流速を測定したところ,切断時刻の範囲においてはインタクト個体の流速は午後2時30分が最も速く48cm/h,午後8時30分が最も遅く9cm/hであった。茎を切断すると流速は急激に低下し,10分後には前者で18cm/h,後者で6cm/hとなり,時間の経過とともに緩やかに低下する傾向を示した(図7) 。
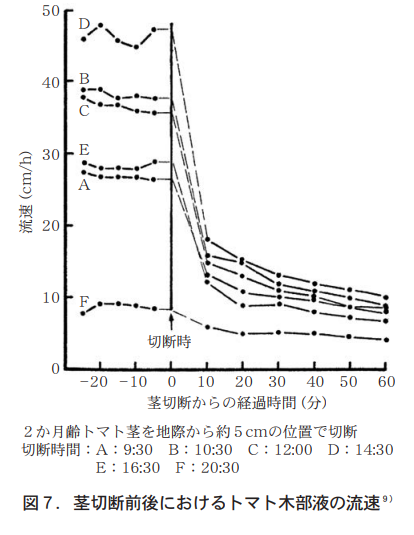
一般に,根から地上部への水の移動速度は蒸散作用と根圧の強度によって律速される。蒸散の盛んな植物では根圧の影響は極めて小さくほとんど無視できる程度で,しかもその根圧は根から離れるにつれて小さくなり,ついには全く見られなくなるとされる18)。従って,この結果は,地上部10cmの部位における蒸散力と根圧の影響の比率を示すものとして理解される。茎を切断すると蒸散力がなくなるため流速は急激に低下し,その値は日中には切断前の約1/4にもなる。このことから,日中の水の移動にはこの部位で蒸散力が約75%,根圧が約25%の割合で働いているものと推定できる。
この時,いっ泌液の成分濃度はインタクト植物の木部液の成分濃度を反映している必要がある。この点については多くの研究者により議論され,ArmstrongとKirkby1)は茎切断後15分〜60分の濃度が安定しており,インタクト個体の濃度を代表させるのに最も良いとしている。筆者らも茎切除後,15分間毎に1時間いっ泌液を採取し分析したところ,切除後1時間内の成分濃度にほとんど変化が認められないことから,この値はインタクト個体の木部液濃度を反映しているものとした9)。しかし,それでもなお養分の移行を動的に把握するためには,生育中のトマト茎に差し込み木部液の成分濃度を検知するマイクロセンサーの開発等が望まれる。
5.昼夜間の木部いっ泌液濃度と培養液濃度の関連性
水,養分が滞留なく根から地上部に動いていると仮定すれば,いっ泌液濃度≒吸収濃度(n/w)となる。勿論,これらを経時的にみれば,前者は点的分析であり後者は線的分析であるので数値が一致することはまずないが,午前0時と正午のいっ泌液の分析値は夜間の,あるいは昼間のn/w値に遠からず近似するものと考えられる。また,根の養水分吸収は苗齢によっても変化することはよく知られた事実である。
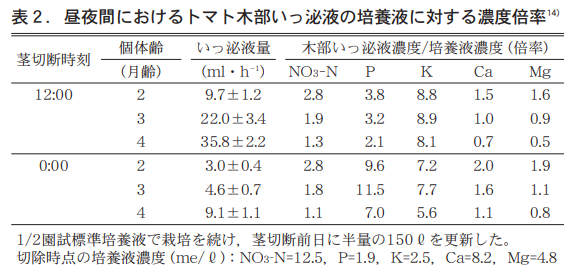
そこで1,2,3月にトマトを播種し園試処方1/2培養液で栽培を続け5月に茎切除し(12:00と0:00)1時間採取のいっ泌液とその時の培養液の分析値から,いっ泌液濃縮係数(XylemSapConcentration Factor (XSCF) を求めた(表2) 。XSCFは,いっ泌液濃度を培養液濃度で除した値で,これはRussel and Shorrocks20)のTranspiration stream concentration factor(TSCF) に準じて筆者が記した名称である9)。
これを見ると,齢が進み樹の大きくなるほどいっ泌液量は多く,いずれの齢でも12:00は0:00の3〜5倍となる。養分のNO3−N,Ca,Mgは齢が進むにつれてXSCFが低くなる。特に4か月齢のトマトではCaで昼0.7,Mgで昼0.5,夜0.6と1以下となり,いっ泌液濃度は培養液濃度より低くなることが分かる。
これに対し,PのXSCFは4か月齢でも昼2.1,夜7.0と高く,かつ夜の方がはるかに高い値を示す。逆にKのXSCFは昼8.1,夜5.6と昼の方が高い値を示す。なお,KのXSCFが昼夜とも非常に高い数値となるのは,表2欄下に記したように分析時の培養液濃度(2.5me/ℓ)が当初の濃度の1/3相当に低下していたことによると考えられる。
以上,論述してきたように,各成分のn/wは吸水量の少ない夜に高く吸水量の多い昼に低くなる。特に夜間のPのn/wは昼間のそれの4〜5倍で他の成分の値よりもはるかに高い。なぜ,Pは水移動の少ない夜間にも他の成分に比べて多く吸収されるのか? Pの多面的な生理機能のうち,夜間にも恒常的に行われる細胞の分裂と増殖,それにかかる核酸および細胞膜の形成,呼吸に必要なATP生産等に関わり,昼間と同等に要求されるからではないかと思われる。
一方,Kのn/wは夜間の方が高いにも関わらずXSCFは昼間の方が高い。夜間の養分吸収とその移行にタイムラグが存在するにしても養分の中でKだけがそうなるとは考えにくい。Ben Zioniら2)は,植物におけるKの地上部から根へのフィードバックメカニズムを提唱し,KはNO3−N のカウンターイオンとして働いているとしている。トマトにおいても地上部から根へ移行してきたKが再び地上部へ移行していることが確認され,それは生育条件によっても異なるが,最も高い時で輸送濃度の20%を占めるとされる1, 7)。筆者も木部いっ泌液のK濃度は明け方から上昇することを明らかにしているが14),これには根に転流してきたKの再転流が関係している可能性が高い。これまで述べてきたように,夜間におけるPの動きと昼間におけるKの動きを32Pと40K(製造するのが極めて困難とされている)で可視化することができれば,それは作物の栽培管理に極めて有用な知見を提供することになる。
6.最後に−今後の展望−
植物によるカチオンの吸収のme総和(NH4⁺+K⁺+Ca2⁺+Mg2⁺+Na⁺)がアニオンの吸収の総和(NO3⁻+SO42⁻+H2PO4⁻+Cl⁻)を超えると,植物は根圏へカチオン(H⁺)を排出しなければならず,その結果,細胞内のpHが高くなるのでホメオスタシス(恒常性)を維持するため,取り込まれた過剰のカチオンは有機酸の合成を高めて細胞内のpHの高まりを調整するとされる16)。生物のもつこのような生理機能も「動的平衡」 3)の一例として理解できる。木部液においてもカチオン−アニオンのバランスは維持されているはずで,夜Pと昼KのXSCFに関連するカウンターイオンは何か,根へ再転流して互いにその一端を担っているのか,大変興味ある問題である。
一方,根の水透過性は日中に高く夜間に低い日周変動を示し,これにはアクアポリン発現量が密接にかかわっているとされる。アクアポリン発現量は暗期開始数時間後に最低となり,その後,明期開始に向けて徐々に増加することから,この発現量の日変動はサーカディアンリズムに支配されると考えられている8)。根特異的なアクアポリン遺伝子が明期開始後に急激に発現するが,地上部の湿度を100%近くに保つとアクアポリン遺伝子の発現誘導が見られなかったことから,根における急激なアクアポリンの発現誘導は,サーカディアンリズムに加えて地上部からの蒸散要求が大きく関わっているとされる21)。
筆者らは,接ぎ木キュウリと自根キュウリについて,木部いっ泌液を48時間にわたり1時間毎フラクションコレクターで受け,いっ泌液量と無機成分を分析したところ,カボチャの根でもキュウリの根でも,いっ泌液量は明らかにサーカディアンリズムを示した。それは午前8時ころから急激に上昇し午後2時にピークとなる。その後,徐々になだらかな曲線をえがいて日出頃に最低となる(図8) 。この時,無機成分のNO3−N濃度は急激に低下し,6時間後には直後の約1/2となるのに対して, P濃度は急激に上昇し6時間後には直後の2倍近くとなった11)。この実験では地上部を除去しているので蒸散の影響は無視でき,正に根そのものの反応(根圧)と理解される。
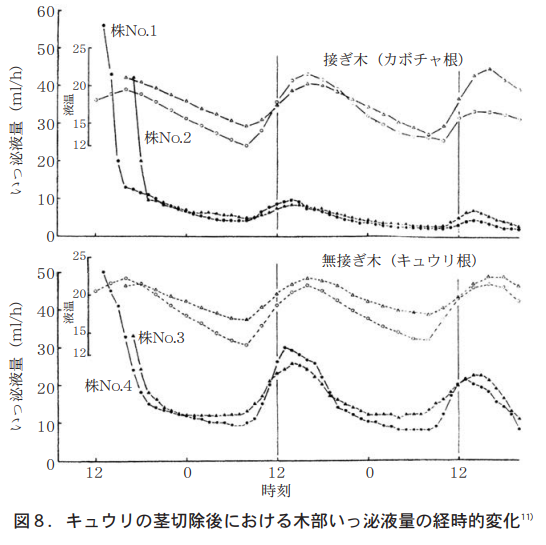
このサーカディアンリズムにはアクアポリン遺伝子はどうかかわっているのだろうか。イネの根では多くのアクアポリン分子種が明期開始数時間後に遺伝子発現量のピークを迎える日周変動を示し,タンパク量ではさらに2〜3時間後にピークを迎える21)。最近,岡山県のノートルダム女子高校の前田彩花さんは,温度一定の明暗条件におけるアジアイネの吸水の日周変動を詳細に調べ,吸水上昇は明期開始前から顕著にみられ,アクアポリンの転写量は明期開始前後に増加することを明らかにし,吸水が蒸散よりも先に発動する可能性の高いことをJSEC−2019(朝日新聞社などが主催する学生を対象とする学術賞「高校生・高専生科学技術チャレンジ」)で発表している。
木部での水輸送が蒸散力と根圧によることはよく知られているが,根圧による水吸収とアクアポリンの遺伝子発現の関連性については未だ報告はみられない。根の吸水には溶液の浸透圧が関係し,加えて養分吸収の促進あるいは阻害を伴なうので,根の機能解析には水と養分を両極に据える必要がある。さらに根は地上部からのシグナルに応答していることを考えると,上述の重水トレーサーやPETIS技術などを個体における動態解析の手段として駆使し,その成果を高い階層レベルの統合的科学6)へ,とりわけ作物栽培・生産学の領域に応用していくことが重要であると言えよう。
引 用 文 献
1.Armstrong, M. J. and E. A. Kirkby. 1979.
Estimation of potassium recirculation in tomato plants by comparison of rates of potassium and calcium accumulation in the tops with their fluxes in the xylem stream. Plant Physiol. 63: 1143-1148.
2.Ben Zioni, A., Vaddia, Y. and S. H. Lips.1971.
Nitrate uptake by roots as regulated by nitrate reduction products of the shoot. Physiol. Plant. 24: 288-290.
3.福岡伸一.2017.
新版動的平衡.小学館新書.−動的平衡とは何か−.260-263p.
4.今井俊治・岩尾憲三・藤原多見夫.1991.
ブドウの生体情報の測定と解析による土壌水分管理法の指標化.3.
土壌乾燥と茎径並びに果粒肥大の日変化特性.生物環境調節29: 19-26.
5.金井俊輔.2009.
トマトの物質生産に及ぼす環境ストレスの影響と
その生体情報に基づく栄養生理学的解析.広島大学学位論文.115p.
6.菊池卓郎.2009.
新しい統合科学としての園芸科学の確立をめざして.
農業および園芸.84:319-326.
7.Kirkby, E. A., Armstrong, M. J. and J. E. Legett. 1981.
Potassium recirculation in tomato plants in relation to potassium supply.
J. Plant Nurti. 3: 955-966.
8.Lopez, F., Bousser, A., Sissoëff, I., Gaspa M., Lachaise, B., Hoarau, J. and A.
Mahé. Diurnal regulation of water transport and aquaporin gene expression
in maize roots: contribution of PIP2 proteins. Plant Cell Physiol. 44: 1384-1395.
9.桝田正治.1989.
トマトおよびキュウリの真昼と真夜中における木部いっ泌液の無機成分濃度.
園芸学会誌.58: 691-625
10.桝田正治.1992.
トマトの夜間における生育と窒素吸収.
平成3年度科学研究報告書(一般研究C) .課題番号02660033: 3p.
11.Masuda, M. and K, Gomi. 1982.
Diurnal changes of the exudation rate and the mineral concentration
in xylem sap after decapitation of grafted and non-grafted cucumbers.
J. Japan. Soc. Hort. Sci. 51:293-298.
12.桝田正治・今野裕光.2021.
トマト,メロンの紐栽培における肥料袋の投入箇所と培地の太陽熱消毒について.
農業と科学728: 1-7.
13.桝田正治・野村眞史.1995.
トマトの摘心及び果実除去が根の養分吸収と酸素消費に及ぼす影響.
園芸学会誌.64: 73-78.
14.桝田正治・島田吉裕.1993.
トマト木部いっ泌液における無機成分濃度の日変化およびその濃度に及ぼす光照度と苗齢の影響.
園芸学会誌.61: 839-845.
15.Masuda, M., Tanaka,T. and S. Matsunari.
Uptake of water and minerals during day and night in tomato and cucumber plants.
J. Japan. Soc. Hort. Sci. 58: 951-957.
16.松本英明.1991.根圏のpHに及ぼす植物の影響.
土壌肥料学会誌.62: 563-572.
17.太田勝己.1996.
ミニトマトにおける裂果発生の機構解明とその制御に関する研究.
京都大学学位論文,52p.
18.Pate, J. S. 1962.
Root-exdation studies on the exchange of 14C-labelled organic
substances between the roots and shoot of the nodulated legume.
Plant and Soil. 17:333 -346.
19.Rudich, J., Rendon-Poblete, E. and A. Abdel-Ilah. 1981.
Use of leaf water potential to determine water stress in field-grown tomato plants.
J. Amer. Soc. Hort. Sci. 106:732-736.
20.Russell, R. S. and V. M. Shorrocks. 1959.
The relationship between transpiration and the absorption of inorganic ions by intact plants.
J. Exp. Bot. 10: 301-316.
21.Sakurai-Ishikawa, J., Murai-Hatano, M., Hayashi, H., Ahamed, A., Fukushi, K.,
Matsumoto, T. and Y. Kitagawa. 2011.
Transpiration from shoots triggers diurnal changes in root aquaporin expression.
Plant Cell Environ. 34: 1150-1163.
22.Terabayashi, S., Takii, K. and N. Namiki.
Variation in diurnal uptake of water and nutrients by tomato plants
of different growth stages grown in water culture. J.Japan. Soc. Hort. Sci. 59: 751-755.
23.塚本崇志・石井里美・七夕小百合・鈴井伸郎・河地有木・藤巻秀・草川知行.2020.
Positron-emitting Tracer Imaging System
(PETIS) 法を用いたトマト果実への光合成産物の転流に果実直下の側枝葉が及ぼす影響の評価.
園芸学研究.19: 269-173.
24.山崎肯哉・鈴木芳夫・篠原 温.1976.
そ菜の養液栽培(水耕)に関する研究.
特に培養液管理と見かけの吸収濃度(n/w)について.東教大農紀要. 22: 53-100.
25.Went, F. W. 1944.
Plant growth under controlled conditions (II). Amer. J. Bot. 31:135-150.
土のはなし−第12回
堆肥の効果の現れ方と土の条件
−土の黒さが決め手−
ジェイカムアグリ株式会社
北海道支店 技術顧問
松中 照夫
化学肥料が世に現れる前の時代,農地に養分を補給するという難題を堆肥という資材を考え出して乗り切った歴史を,前回,振り返った。またヨーロッパの畑とわが国の水田では,農業のあり方がちがい,それが堆肥に対する考え方のちがいを生んだことをお話しした。化学肥料が世に出た後も,堆肥には多様な効果が期待されている。しかし,その多様な効果が例外なくすべての土で発現するのではない。その効果の現れ方のちがいをもたらす土の条件は何か,それを考えてみる。
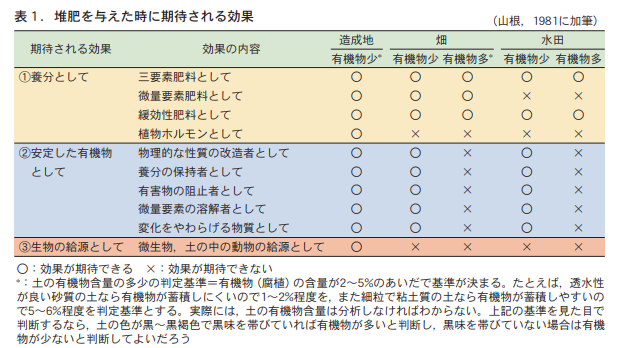
1.堆肥に期待される効果
堆肥には,大きく分けて3つの効果が期待されている (表1) 。①養分としての効果,②比較的分解されにくい安定した有機物としての効果,③生物の供給源としての効果である。堆肥を与えれば,この3つの効果が同時に自動的に現れるように,しばしばいわれる。堆肥を与えると土がよくなると思われるのはそのためだろう。しかし,この3つの効果は「期待される効果」であって,必ずその効果が現れるわけではない。
1)養分としての効果
まずは,養分としての効果である。堆肥を与えることで直接的に期待できる効果は,この効果である。具体的には,①多量要素,とりわけ三要素(窒素,リン,カリウム)の供給源,②微量要素の供給源,③ゆっくりと効果があらわれる肥料(緩効性肥料)としての効果,④植物ホルモンの供給源,などである。これらの効果のうち,土の条件にかかわらず,堆肥を与えることで効果が確実に期待できるのは,①の三要素肥料としての効果である。通常の畑や水田の土で,窒素,リン,カリウムのいずれもが作物生産の制限因子とならないという土は考えにくいからである。また,③緩効性肥料としての働きも,土の条件にかかわらず期待できる。それは,堆肥が土に与えられた後,土の中の動物(トビムシ,ワラジムシ,ミミズなど)や微生物(細菌,放線菌,糸状菌など)などが協力して堆肥を分解し,その分解にともなって堆肥から養分が徐々に放出されるからである。堆肥を連用すれば,累積的で持続的な養分効果も期待できる。
ところが,②の微量要素肥料としての効果は,水田ではあまり期待できない。これは,水田のかんがい水中に微量要素がかなり溶け込んでおり,イネ栽培期間に多量にとりこまれるかんがい水から供給される微量要素量が多いためである。堆肥に微量要素が含まれていても,その与える量が限られた範囲では,かんがい水からの天然供給量にはおよばない。
④の植物ホルモンも,栽培履歴のある畑や水田で,実際にどれほどの効果が期待できるかはまだよく知られていない。ただし,造成地のように,有機物を含む表層土が完全に除去され,有機物をほとんど含まない下層土が作土になっている場合には効果が期待できそうである。
2)安定した有機物としての効果
期待できる効果の二つ目は,安定した有機物としての効果である。安定した有機物とは,土の中で動物や微生物などの分解をある程度受けた後に残った有機物で,比較的分解しにくい有機物のことである。これが土の有機物(腐植)といわれる物質となり、土に黒い色を与える。
堆肥が土の中にすき込まれると,堆肥の中の分解されやすい有機物は,分解されることで養分的効果を現す。一方,比較的分解されにくいために土に残った有機物は,土の中にもともとあった有機物とともに安定した有機物としての効果を発現する。その効果には,①土の物理的な性質,たとえば土のすき間の大きさやその割合(孔隙分布) ,排水のしやすさ(透水性) ,水持ちのよさ(保水性) ,空気の通りやすさ(通気性) ,耕しやすさ(易耕性)などの改良,②養分保持能の増加,③有害物の抑制,たとえば,有機物がアルミニウムと結合すると,アルミニウムの有害な働きを抑えるため(キレート作用という),アルミニウムがリンと結合しにくくなる。
その結果,リンの養分効果が出やすくなるといった効果,④微量要素は水に溶けにくい形態であることが多い。しかし,有機物が分解されるにともない二酸化炭素(CO2)が放出され,これが水に溶けて炭酸水となって微量要素を溶けやすくする働き,さらに⑤有機物の持つ環境変化をやわらげる作用(緩衝力)などが考えられる。
しかし,こうした様々に期待される安定した有機物としての堆肥の効果が現れるのは,与えられた土の有機物含量が,ある基準(土によってちがい,2〜5%くらいの範囲)より少ない場合で,それ以上の場合には効果が期待できない(山根,1981) 。有機物が多い土では,もともと土に安定した有機物(腐植)が多いので,土の物理的な性質が作物生産の制限因子になりにくいからである。
3)生物の供給源としての効果
三つ目の効果は生物の給源としての効果である。堆肥中には多くの生物(ミミズなどの小動物や微生物など)が生息している。堆肥を与えることは,土の中にこれらの生物を供給することになるので,その供給源としての効果が期待できる。しかし,この効果も堆肥を与える土が通常の土であれば,その土に生息する生物数が,与えられた堆肥に含まれている生物数にくらべて圧倒的に多く,堆肥に土の生物の給源としての直接的な効果を期待しにくい。この効果も造成地のような極度に有機物の少ない土が作土となった場合に限定すべきである。
堆肥を与えることが土の生物におよぼす影響は,1年間の効果としてとらえるよりも,連用による累積的な効果のほうが期待できる。ただし,その場合でも,生物数の多様化や増加が作物の生育に直接的な影響を与えるかどうかは,そのほかの土の条件によって変化する可能性がある。
2.有機物の少ない土ほど堆肥の効果は大きい
今,土に堆肥を与えようとする場合,とくに理由もなく,ただ漫然と「土づくり」だからといって与えていないだろうか。なぜ堆肥を与えるのか,堆肥のどの効果を期待して土に与えるのか,それをよく考える必要がある。土の条件によっては,堆肥に期待した効果が現れる場合と,現れない場合とがあるからだ。表1に示したように,土の有機物が多いか少ないかが判断基準となる。
土の有機物の多少は,厳密には分析しなければわからない。土を分析することなく見た目で有機物の多少を判断するなら,土の色を見るとよい。土の色が黒〜黒褐色で黒い色を帯びていれば有機物が多いと判断し,土の色が黒い色を帯びていない場合は,有機物が少ないと判断して大きなまちがいはないだろう。
有機物の少ない土(黒い色が淡い土)は堆肥を与えることで多様な効果が期待できる。有機物の多い土(黒い色が濃い土)では,緩効的な三要素肥料としての養分効果を中心に期待したい。