
第760号 2024(R06).05発行
PDF版はこちら
§園芸作物の光応答反応と農業技術(2)
-その他,花・野菜への光照射と花成制御の理論的背景-
元 岡山大学大学院自然科学研究科
桝田 正治
§愛知県における農作物鳥獣被害防止対策の取組
愛知県農業水産局農政部農業振興課
野生イノシシ対策室
辻井 修
§土のはなし-第31回
危機に瀕する世界の土-その1
古代文明の崩壊と土の劣化
前 ジェイカムアグリ株式会社
北海道支店 技術顧問
松中 照夫
園芸作物の光応答反応と農業技術(2)
-その他,花・野菜への光照射と花成制御の理論的背景-
元 岡山大学大学院自然科学研究科
桝田 正治
1.園芸作物の電照
1)花卉類
日長制御による開花促進は,多くの長日性の花卉類で明らかにされており,光源も白熱電球,蛍光灯からLEDに代わりつつある。代表的な切り花としてシュッコンカスミソウ,ストック,トルコギキョウ,キンギョソウがあげられる。シュッコンカスミソウは,フラワーアレンジメント,いわゆる添え花として古くからその独自性を発揮してきた。その花成は赤色光では促進されず遠赤色光〈これに少し赤色光が加わると効果大〉によって促進され,多くの県で周年栽培が可能となっている。遠赤色光の効果はストックでも認められる1)。ストックの需要は年末から3月とされ,千葉県は全国一位で約10億の生産額を誇る。アイアン系の1本立ち主力品種では9月中旬から16時間日長となるよう遠赤色LEDの電照が開始される。トルコギキョウも,遠赤色光によって花成と茎伸長が促進される2)。熊本県の冬出し栽培においては花蕾のブラスチング〈よく発達した花芽が枯死する現象〉も抑えられるといった報告もみられる。赤色光にはほとんど効果は見られない。ただ,定植時から摘心時までの赤色LEDに切り花の品質向上効果が認められ,千葉県や埼玉県では摘心前の赤色 LED+摘心後の遠赤色LED(いずれも終夜照明) で秋出しの作型が定着しているようである3,4)。キンギョソウも遠赤色LEDに反応し,摘心時より日長延長(16時間)あるいは3時間の暗期中断が一般に行われる5)。以上のように,相対的長日性の作物では遠赤色LEDの電照が開花を促進し,切り花生産の安定化に資する技法となっていることが分かる。
2)野菜類
花卉に比べ野菜の電照栽培は少なく,花芽を抑制し葉を商品とするシソぐらいである。典型的な短日植物であるシソは暗期中断1時間でも花芽抑制効果の高いことが既に30年前に宮崎県において報告されている6)。ホウレンソウは長日植物であるが,成松7)は秋冬期に白熱灯で日長13時間程度にすれば花成に影響せずボリュームの増大がはかれることを示した。近年,山崎8)は,赤色LEDの2時間照明により品種‘ミラージュ’の葉面積が大きくなること,明け方の照明は真夜中の照明と比べて花茎伸長を抑制するとした。この報告は栄養成長を促す暗期中断の時刻について,暗期の中心以降に光照射の技術が存在すること,同時に電照効果は長日に鈍感な晩抽性の品種でより強く発現する可能性を示唆するもので,先に述べたキク暗期中断の時刻(Max)を念頭に秋冬期ハウスホウレンソウの草勢強化を図ることは,とりわけ生育緩慢な70%シュウ酸低減の新品種‘まろみ’ (2022年発売,トーホク種苗)には必要ではないかと筆者は考えている(図1)。

2.紫外光および黄色LEDの照射
1)UV-B照射によるイチゴのうどん粉病とハダニの抑止効果
紫外線(UV-A:315~400nm)は,地表に到達する紫外光のほとんどを占め,ナスやプリムラマラコイデスの色素生成に必要なことは,古くから知られている。例えば,ナスの色素ナスニンは 370nm付近の光が必要とされ,それを遮断するガラス室での栽培では色付きが悪くなる。また, UV-B(280~315nm)は,バラのうどん粉病やハダニを抑制するが,イチゴでも紫外線の照射効果は高く,農研機構中央農業研究センターは多くの研究機関を統括して6年間の研究成果を2019年にイチゴ病害防除マニュアルとして公表した9)。導入にあたって特に重要と思われる点は,ハダニ抑止効果としてUV-B照射は3時間でよく,照射後に太陽光に当たると卵のふ化率が4時間のうちに 50%回復することから,UV-B照射は少なくとも日の出前3~4時間に終えること。さらに葉裏の光を増強するために光反射シートを併用するとある。その他,天敵カブリダニや植物賦活剤の併用についても記述されているが,研究代表者の後藤千枝氏は農薬について70%以上削減できる防除体系を構築することができたと冒頭で総括している。このことからも分かるように,本研究の成果は,いわゆる IPM(Integrated Pest Management), 「総合的病害虫管理」の概念に位置付けられコストパフォーマンスの向上に資する技術と言えよう。
2)UV-C照射による黄ニラの緑化抑止効果
岡山県農林水産総合センター農業研究所の岡氏 (野菜・花研究室長)によれば,天日干しの黄ニラでも市場に出すとどうしても弱光に当たるので徐々に緑色を帯びてくるという。このニラにあらかじめUV-C光(200~280nm)を照度5Klxで4時間照射すると少なくとも40時間は全く緑化しない10)(図2)。この反応を見出した時点でこの技術はすでに他社によって特許公開(特開2013- 48615)されていたが,未審査請求による見なし取り下げ〈平成26,11,25〉になっているとのこと。このUV-Cがクロロフィル生成のどの段階をブロックするのか,筆者の知る限りではその機作は今なお明らかとはなっていないが,核酸の吸収スペクトルのピークがUV-C波長域内にあるので,光が細胞の核酸にダメージを与えクロロフィルだけでなく,あらゆる代謝機能を阻害しているものと考えられる。

3)点滅型黄色LED照射によるキクの害虫忌避効果
私の住んでいる赤磐市の桃園一体は,6月になるとヤガ忌避のため黄緑色の防蛾灯がともる。桃は幼果で,このとき光の影響は全く問題とされない。しかし,日長を感受する作物では問題となる。特にキクでは花の発育に影響するため夜間照明はできない。キク栽培における「開花遅延の回避」と「防蛾効果の発現」を同時に解決するため,石倉11)はパルス光に着目し,キクの発蕾に及ぼす影響を詳細に調べ黄色LEDパルス光の有用性を示した。そして,2018年に点滅形LEDランプを製品化し,トマトやホウレンソウにも適用できることを現地圃場において検証し,当製品の普及には研究員,普及員の現地との連携による対人ネットワークの人材活用が必要であるとした12)。研究成果を広く普及するには関係者による情報の交換と相互の議論,それらを通じて現地ニーズとの共有化をはかることの重要性が示唆されている。
3.日長(電照)による花芽分化にかかる生体内反応
本稿で取り上げたキクの電照は弱光によって花成を制御する技術で,この引き金的な役割を果たすのは,赤・遠赤色光受容体のフィトクロム (phytochrome)であると考えられる。これは分子量約11万の水溶性蛋白質で,図3に示したようにPr型(吸収極大660nm)とPfr型(吸収極大 730nm)で存在し,表記の波長を中心とする光照射で相互に変換するといった特性をもつ。暗所で合成されたPr型フィトクロムは主に細胞質に分布するが,Pfr型に変換されると核内に移行しシグナル因子と相互に作用して特定の遺伝子の発現を調節するとされる。フィトクロムはA, B, Cの3つのグループに大別され,なかでもAは最も光感受性が高く,Pfr型に変換されると暗所で速やかに分解される特性を有する。キクの暗期中断におけるPr型,Pfr型の葉内での動きを,まず図3のフィトクロムの特性に照らして解説すれば以下のようになる。
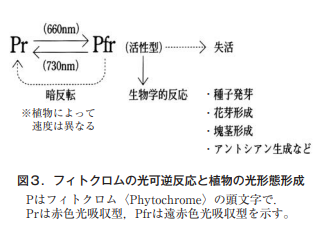
短日植物のオナモミやアサガオは,正確にいえば長夜植物である。長い夜によって花成が誘導されるのである。アサガオの品種‘紫’は16時間の暗期を1回与えると花芽を形成するが,葉で作られた花成刺激が芽に移動する速度は巧妙な実験により51cm/時間と計算された13)。フィトクロム説では,16時間という長い夜の間にPfr型フィトクロムは,Pr型に徐々に変換していき〈図中の暗反転〉,この活性型Pfrがある限界量以下になると花成が誘導されるというものである。したがって,これが8時間のような短い夜では短日植物は, Pfrが限界量以下とはならないため花成は誘導されないことになる。
われわれの生活に身近なキクの花成も同様にこの可逆反応により主に制御されていると考えられるが,赤色光で暗期中断を行うとPrがPfrに変換し,Pfrが限界量を超えるので花成は抑えられることになる。ただ,実際には照射時間が3~5時間と長く,赤色に遠赤色を加えても電照効果が変わらない,むしろ効果大とする報告が多く見られることは,図中に記載したPfrの分解による失活が関係するものと推察される。特にフィトクロムAの量的安定性を保つには,照射時間が長い場合に遠赤色で一部Pr型に変換することにも意味があると考えれば,これも種の生存に向けた光受容体フィトクロムの動的平衡として理解できる。この理由を白山(前報の文献3参照)はフィトクロム作用特性から,キクではフィトクロムAと遠赤色による高照射(エネルギー量が高くなる?)反応が関与している可能性を指摘し,詳細な機構の解明は今後の課題であると記している。
葉で作られた活性型の蛋白質Pfrは,そのあとの花成反応にどのように受け継がれるのであろうか?近年,遺伝子レベルで研究が進み,FT遺伝子〈Flowering locus T〉によるFTタンパク質がシロイヌナズナで発見され14),この遺伝子は2005 年に花成ホルモン「フロリゲン」と命名された。その後,葉から茎頂に移動したこのFTタンパク質は茎頂に存在するFDタンパク質と一緒に働いて花芽形成遺伝子〈AP1遺伝子〉をオンにすることが示され,そのFT-FD複合体を作る過程が阿部らによって可視化された15)。
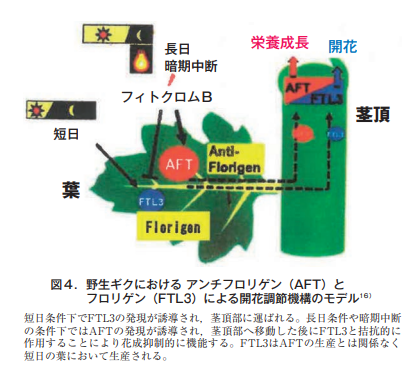
一方,キクの柳芽に代表されるように花成の非誘導条件に〈一部の葉でも〉移されると栄養成長に移行する現象はキクだけでなく多くの植物で示され,葉における花成抑制物質の存在が示唆されてきた。樋口ら16)は,2倍体野生キクのFT遺伝子には3種類〈CsFTL1, 2, 3〉あること,このうちFTL3は花成促進に強く働くが,FTL3に拮抗的に作用するAFT遺伝子〈AFT: Anti-florigenic FT/TFL1 family protein〉を発見し,それをアンチフロリゲンと称した。花成の非誘導条件下の葉〈長日や暗期中断〉で作られるAFTは茎頂に移動して花成を抑制する(図4)。長日から短日に移されるとAFTレベルは急激に減少する一方,FTL3は短日周期が繰り返されることによって徐々に誘導される(図4)。ここでは暗期中断を感知する主要な光受容体としてフィトクロムBがアンチフロリゲン(AFT)の発現を誘導する一方で,フロリゲン(FTL3)の発現を抑制するとしている。ところが,短日下のFTL3の誘導にはフィトクロムBの関与は示されていない。フィトクロムは赤色・遠赤色可逆の色素タンパク質で常に葉に存在しているので,短日でもこのシステムは作動しているはずである。少なくとも最初の短日光周期サイクルではそう言えるであろう。短日下のフィトクロムBの関与が遺伝子レベルで明らかになれば,光レセプターとしてのフィトクロムからFT(FTL3)とAFTへの研究の流れ〈エネルギーの受け渡し〉は,私にはよく理解できるのである。
花成研究の歴史は長く,今や茎頂における遺伝子解析の段階にあるといえる。茎頂分裂組織での質的変換に関係する一連の遺伝子(タンパク質)は,今後も多々発見されるであろうが,それらを農業に利用するためには花器形成まで含めて花芽促進作用を持つタンパク質FTL3様物質の周辺が広く埋まることではないかと思料される。
おわりに
地球にふり注ぐ太陽光の放射エネルギー比と植物に有効な波長域を図5に示した。波長700nm以上の遠赤色は光合成には有効でないが,本論で述べてきた形態形成には極めて重要な光となることが分かる。この波長の光は薄暗い。800nmにもなると目では見えない熱線となる。植物の光応答は,炭酸固定にみられる高エネルギー反応と形態形成にみられる低エネルギー反応に大別される。
前者は,いわゆる光合成で6CO2+12H2O*+ (光エネルギー)→ C6H12O6+6O*2+6H2Oの化学式で示されるように光エネルギーを使って炭酸ガスと水から糖を合成する反応である。イモ類では根や茎にデンプンとして貯蔵され,果実では蔗糖や果糖として集積される。糖の一部はC6H12O6+ 6O2 → 6CO2+6H2O+化学エネルギーで示されるように,生存に必要なエネルギーを産出している。この反応が呼吸で,植物は酸素を使って炭酸ガスと水を放出しているのである。光合成研究は,岡山大学の沈教授が世界をリードし,水分解反応の機構と酸素の放出に関する画期的な成果を発表されており,理学部のホームページには太陽光エネルギーを利用して水分解反応を人工的に行う「人工光合成」のための触媒創成に重要な基礎を提供するものである,と記されている。
本稿では,後者の低エネルギー反応,とりわけ電照を取り上げたが,一概に電照と言っても,上述してきたようにキクでは花芽分化への質的変換を,イチゴでは矮化抑止への量的拡大を意味し,その目的はキクとイチゴで異なる。花芽形成を促進する点では両者とも夏から秋への短日(長夜) が働く。キクでは電照により花芽形成が抑制されるが,イチゴでは低温が主要因となって働いているため電照しても花芽形成は続く。ただし,電照時間が長すぎると,つまり長日に過ぎると花芽分化に支障をきたす。イチゴの葉身や葉柄の伸長を促す電照は生育の具合を見て修正する必要があり,キクの電照よりもきめ細やかな技術手法が求められる。葉で感受した短日刺激は茎頂に伝わる。キクについては,電照や暗期中断によってAFT が強く働きFT(FTL3)遺伝子は抑止され花芽分化は起こらない。一般に植物は,ある一定の齢 (相)を経なければ日長を感受して花成を創始することができない。この相を「幼若相」と称するが,この相ではまた別の遺伝子が働いているのであろうか?大変興味ある点であり,また別の機会に論じたい。
イチゴにおいても電照による葉での長日刺激は茎頂に運ばれているはずで,それは少なくとも葉の量的拡大に作用している。イチゴでは植物ホルモンのひとつであるジベレリンの消長が認められているが,葉から送られたタンパク質が茎頂でのジベレリン合成に関係している可能性は多分にある。イチゴの葉で作られるタンパク質が,花成反応とは別に葉の伸長成長にかかわる働きを有していることは確 かで,電照によるイチゴの栄養成長にAFT遺伝子は如何にかかわっているのであろうか?はたして8倍体の栽培イチゴにも AFT様,FT様の遺伝子がキクと同様な機構で存在するのであろうか?ただ,イチゴの花芽分化には,前報の図9に示したように低温が主導的役割を果たすので,フロリゲン〈日長で誘導される花成ホルモン〉とは異なる仮称未確認バーナリン〈低温で誘導される花成ホルモン〉タンパク質の存在も予想され,一連の研究が農業的価値の高いイチゴ電照の場にも新たな光を灯してくれる,その日の来るのが待たれる。
引用文献・資料
1.種谷光泰・加藤美紀・椎木千晴・香川晴彦.2016.
遠赤色光照射によるアイアン系ストックの開花促進技術.
千葉農林総合研研報8:41-50.
2.工藤陽史・山口 茂・佐藤 旭・栗山孝浩・深井誠一.2012.
熊本県のトルコギキョウ冬出し栽培における電照が開花と花蕾のブラスチングおよび茎伸長に及ぼす影響.
園芸学研究11(3):363-369.
3.千葉県農林総合センター.2019.
赤色LEDと遠赤色LED光源を組み合わせた秋出しトルコギキョウの品質向上.
試験研究成果普及情報.
4.さいたまの花普及促進協議会.2022.
トルコキキョウにおける11月出荷高品質栽培管理技術.
5.千葉県農林総合センター.2019.
キンギョソウ新品種の特性と電照及び据え置き栽培法.
試験研究成果普及情報,
6.飯田久雄・細山田芳博・高橋英生.1989.
暗期中断による大葉シソの抽苔防止.
九農研51:204p.
7.成松次郎.1996.
ホウレンソウの補光栽培に関する研究.
神奈川県農業総合研報137:17-23.
8.山崎基嘉.2019.
チューブ型赤色LEDを用いた電照による
施設ホウレンソウの栽培期間短縮化条件の探索.
新近畿中四国農業研究2:46-54.
9.農研機構中央農業研究センター.2019.
紫外光照射を基幹としたイチゴの病害虫防除マニュアル~技術編~.1-51.
10.岡 修一.2015.
黄ニラの高品質・安定生産技術の確立.(2)緑化抑制技術の確立.
岡山農業総合技術センター 単年度試験研究成績.127-128.
11.石倉 聡.2014.
切り花ギクに利用可能な黄色LEDパルス光を用いた害虫防除技術の開発.
広島総研農技セ研報90:1-88.
12.西濱健太郎・石倉 聡・星野 滋・岡田牧恵・石田真由美.2018.
現地栽培圃場における点滅形LED黄色防蛾ランプの実用性および研究と普及の連携が普及指導員の資質向上に及ぼす影響の検証.
広島総研農技セ研報92:11-22.
13.Wareing,P.E. and Phillips,L.D.J. 1970.
THE CONTROL OF GROWTH & DIFFERENTI-ATION IN PLANTS.
Pergamon Press, Oxford. Chapter10: The physiology of flowering-photoperiodism. 164-195.
14.Kobayashi, Y., Kaya, H., Goto, K., Iwabuchi, M., Araki, T. 1999.
A pair of related genes with antagonistic roles in mediating flowering signals.
Science 286 : 1960-1962.
15.Abe, M., Kobayashi, Y., Yamamoto, S., Daimon, Y., Yamaguchi, A.,
Ikeda Y, Ichinoki, H., Notaguchi, M., Goto, K., Araki, T. 2005.
FD, abZIP protein mediating signals from the floral pathway integrator FT
at the shoot apex. Science 309 : 1052-1056.
16.Higuchi, Y., Narumi, T., Oda, A., Nakano, Y., Sumitomo, K., Fusaki, S., & Hisamatu, T.2013.
The gated induction system of a systemic floral inhibitor, anti-florigen,
determines obligate short-day flowering in chrysanthemums.
Proc. Natl. Acad. Sci. USA 110:17137-17142.
愛知県における農作物鳥獣被害防止対策の取組
愛知県農業水産局農政部農業振興課
野生イノシシ対策室
辻井 修
1.はじめに
農作物の鳥獣被害は全国共通の問題である。市街化が進展し,工業県のイメージが強い愛知県も例外ではない。ここでは,農作物鳥獣被害防止対策の基本と愛知県での取組の概要を紹介する。
2.農作物鳥獣被害防止対策の基本
(1)基本の「三本柱」
農作物鳥獣被害対策では,以下の3つの取組が重要で,いわば「三本柱」である。
① 寄せない:鳥獣が寄りつきづらい環境を保つ
② 入れない:柵や網で侵入を防ぐ
③ 捕まえる:加害個体を捕獲する
いずれも目新しさはないが,21世紀の今日においても他に有効な手立てはない。
(2)寄せない,入れない,捕まえるの順で実施する
「三本柱」は実施順序が重要である。鳥獣対策というとまず捕獲(駆除)を想定しがちだが,捕獲のみ実施してもさしたる効果は得られない。捕獲の前提として,地域の農家・住民による環境管理及び侵入防止対策が必須である。そもそも,農作物を狙う個体をすべて捕獲することは不可能である。農家自身が基本的な対策を実施した上で,捕獲を仕上げに実施するという感覚が望ましい。
(3)捕獲活動は地域ぐるみで
捕獲には原則的に狩猟免許が必要で,特殊な技能が求められる上に危険性もある。さらに近年は,地域の捕獲の担い手不足が深刻化している。地域ぐるみの活動なくして存続は困難になりつつあり,農家・住民の積極的な参画が望まれる。
3.愛知県の取組
(1)愛知県の被害状況について1)
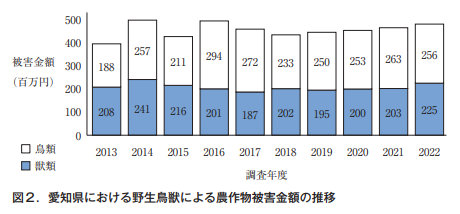
愛知県では,年間4億5千万円前後の農作物被害が発生しており,ほぼ横ばいで推移している (図2)。全国的には被害金額の約80%を獣害(哺乳類)が占めるが,愛知県においては同等か,鳥害の方が多い(図2)。また,園芸作物の被害金額が約65%に達し,野菜や果樹などの園芸作物の生産が盛んな本県農業の特徴を反映していると考えられる。
(2)中山間地域のイノシシ,シカ対策
愛知県中部から東部にかけての中山間地域では,イノシシ,シカによる被害が深刻である。農作物被害のみならず,イノシシの掘り返しによる農地や農道の損壊,交通事故,マダニの蔓延,人身被害に対する不安など,地域全体に影を落とす問題となっている。

ア.集落環境点検と改善活動
前述の「三本柱」を実践するためには,まずは集落の環境を点検し,餌資源と潜み場の解消が求められる2)。最も基本的な対策であるが,愛知県内では十分な取組が行われているとはいえず,残念ながら本稿ではこれといった事例を紹介できない。中山間地域の農村環境保全を図る上で,活動強化を目指さなければならないと考えられる。
イ.侵入防止柵の整備
愛知県内では2010年代以降,国の補助金(鳥獣被害防止総合対策事業)による集落単位でのイノシシ,シカの侵入防止柵整備が進展した。効果を上げるためのポイントとして,以下の3点を提示している。
① 問題となる野生獣に対応可能な仕様とする
② 野生獣の侵入経路をすべて閉鎖する
③ 継続的に点検と補修を行う
それぞれについて解説する。
① 問題となる野生獣に対応可能な仕様とする
シカは跳躍力に優れるため,一定以上の柵の高さを確保する必要がある。筆者らが設置したトレイルカメラで,高さ120cmのワイヤーメッシュ柵をシカが助走なしに跳び越える様子が撮影された (図3)。シカの飛び越え防止の高さは150cmで十分との指摘もあるが3),本県の補助事業の仕様ではシカ対策は高さ180cm以上としている。
サルや中型獣(ハクビシン,アライグマなど)は,ワイヤーメッシュ柵単独での侵入防止は不可能である。これらの獣種に対しては,目合いの細かいネットと電気柵を組み合わせる必要がある。電気柵は電圧を維持するための管理が重要となるが,大区画での共同管理では効果の持続が難しい4)。そのため,まずは個別の農家による対策が望ましい。
② 野生獣の侵入経路をすべて閉鎖する
侵入防止柵は,野生獣の侵入経路になる可能性があるところを極力すべて閉鎖することが重要である。野生獣の侵入経路のパターンは地理的条件や周辺環境によって異なる。侵入経路のパターンに応じ,適切に設置された地区では安定した効果が得られている。
後背に山や丘陵を背負い,山麓に農地や市街地が広がる地区では,山裾沿いの直線的な侵入防止柵設置によっても一定の効果が得られている。イメージとしては,「万里の長城」のような設置方法である。このケースでは,山から流れ出る河川や水路,山林に通じる道路が閉鎖できずに開口部となる。使用頻度が低い道路(林道など)は門扉で閉鎖することで,効果が安定する。河川沿いでは川床に降りるための階段が侵入経路になることがあるので,降り口にワイヤーメッシュを立てるとよい。

周囲を山林に囲まれ,多方向から野生獣が侵入する地区は,農地を完全に囲うことが望ましい。囲い方が不完全だと,回り込まれて容易に侵入を許してしまう。千葉県で実施された調査においても,不完全な囲い方が被害収束を妨げる主因であると指摘されている5)。ただし各筆の田畑を個別に囲うのは非効率でコストが掛かるため,できるだけ多く区画をまとめて囲うことが望ましい。
③ 継続的に点検と補修を行う
柵は設置して終わりではなく,機能を保つための管理が重要である。保守点検状況には集落ごとに差があり,こまめな活動が行われている地区は高い効果を上げている。山端らは獣害対策(本報告ではサルの追い払い活動)の継続により地域のソーシャル・キャピタル(社会関係資本)指標の改善が認められたと報告している6)。侵入防止柵の管理作業もこれと同じく,住民同士のつながりを維持,強化する副次的な効果が期待される。
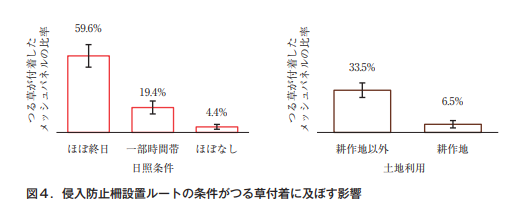
ワイヤーメッシュの侵入防止柵の管理では,つる草の付着に悩まされることが多い。代表的なつる性雑草であるクズは,発芽2年目にはつるが年間17m弱伸長するとの調査事例もあり7),ワイヤーメッシュ柵が完全に呑み込まれる生育量である。筆者らの調査(未発表)において,日当たりがよく陽樹(アカメガシワ,ウルシ類など)が生えているところではつる草の付着が多く,農耕地周辺では比較的少ないことが明らかになった(図4)。日当たり,植生,土地利用等にも配慮することで,効率的な保守点検が可能になると考えられる。
ウ.地域ぐるみの捕獲活動
被害防止対策を目的とした鳥獣捕獲を行うためには市町村長等の許可が必要で,許可申請には原則的に狩猟免許が必要である。そのため,全国的に狩猟者団体構成員が主たる担い手となっており,愛知県も例外ではない。かつて我が国の農山村には狩猟をなりわいとする者がおり,農林業被害対策のための捕獲とのトレードオフで狩猟権を確保していたとの指摘もある8)。
近年,狩猟者団体構成員の減少と高齢化による担い手不足が深刻化している。そこで愛知県では,2009年度の豊根村を皮切りに,狩猟免許を持たない地域住民による捕獲補助活動(有害鳥獣捕獲における狩猟免許を有しない従事者容認事業 (以下「従事者容認事業」という。))9)の普及拡大を図っている。従事者容認事業の導入により,以下のメリットが考えられる。
① 捕獲従事者の作業負担が緩和される。
② わな設置数増加と管理状態の改善により,捕獲実績が伸びる。
③ 従事者容認事業の参加者から,新たな捕獲従事者育成が期待される。
被害対策は当事者が取り組んでこそ,活動が持続する。ちょっとした作業補助であっても,農業者の捕獲活動への参画は意義深い。地域の結束力強化にもつながると考えられ,積極的な取組を促している。
(3)園芸農家のカラス,中型獣対策
愛知県は野菜や果樹など,園芸作物の生産が盛んで,鳥獣被害の報告も多い。特に,カラスなどの鳥類,ハクビシンなどの中型獣による被害が深刻である。愛知県農業総合試験場による県内の果樹農家を対象とした意識調査によると,対策の重要度は鳥類,中型獣,大型獣の順で,中型獣に対しては被害金額の比率以上の脅威を感じていると推察された。また,侵入防止対策を最優先の対策と捉えているとの分析結果が得られた10)。実際に,県の農業普及指導センターには,果樹農家から鳥類及び中型獣被害の相談が多数寄せられており,実用的な技術の現地実証に取り組んでいる。
ア.テグス・ステンレスワイヤによるカラスの侵入防止
農研機構の実証結果11)を参考に,テグス・ステンレスワイヤによる侵入防止対策の果樹園での検証を進めている(図5)。概ね期待どおりの侵入防止効果が得られ,今後の課題は設置・管理作業の省力化である。果樹園は周辺環境が変化に富んでおり,防風林や建物など,設置作業の妨げになる要因も多い。作業体系のちょっとした工夫に加え,テグス展張にドローンを活用するなど,今後の改善が期待される。
なお,本県における効果検証は今のところブドウ,ナシ,モモなど,単位面積当たりの販売額が大きい果樹に限られている。今後は,カキ,露地ミカンなど,より広い面積で栽培される果樹類,キャベツやブロッコリーなどの露地野菜等でも費用対効果が得られる手法の検討が課題である。
イ.中型獣の侵入防止対策
比較的販売単価が高いブドウなどの果樹栽培において,埼玉県が開発した中型獣侵入防止施設「楽落くん」12)の普及拡大を進めている。2017年4月1日以降,狩猟免許の有無に関わらず,農業者自身の事業地における小型はこわなによるハクビシン,アライグマ等の捕獲が許可されるようになったことから13),侵入防止と捕獲を組み合わせた対策を推進しているところである。また,残渣処分の重要性も指摘されているため14),適切な対処を呼びかけている。
(4)野生イノシシの豚熱まん延防止対策
2018年9月に岐阜県で国内26年ぶりとなる養豚農場での豚熱発生が確認され,直後に周辺の野生イノシシの感染も判明した。愛知県では同年12月に,岐阜県境に近い犬山市で豚熱に感染した野生イノシシが見つかった。その後,2023年度末までの累計で193頭の感染個体が確認されており,いまだ終息が見通せない状況である。現在,主に以下の対策を実施している。
ア.野生イノシシの検査
捕獲された野生イノシシから血液等の検体を採取し,検査機関においてリアルタイムPCRによるウイルス遺伝子検査を実施している。一部の検体は抗体検査にも供し,豚熱ウイルスに対する免疫の獲得状況も調査する。結果は関係者に共有し,後述の経口ワクチン及び捕獲強化対策の基礎資料として活用している。
イ.豚熱経口ワクチンの野外散布
野生イノシシに豚熱ウイルスに対する免疫を付与するため,ヨーロッパ諸国での先行事例15)に基づく国の指針に従い,経口ワクチンの野外散布を実施している。経口ワクチンは,弱毒ウイルスを含むワクチン液カプセルをトウモロコシ粉と油脂からなる餌で包んだ製剤で,野生イノシシ生息域の地中に埋めて食べさせる(図6)。
ウ.捕獲強化
捕獲活動経費補助の増額による活動意欲向上,鳥獣保護区や国有林等での認定鳥獣捕獲等事業者による集中的な捕獲活動を実施している。
4.おわりに
愛知県では鳥獣被害防止対策及び野生イノシシの豚熱まん延防止対策について,ウェブサイト及びSNSでも情報発信している(図7)。

5.引用文献,参考資料
(1)愛知県農業水産局.野生鳥獣による農作物被害の状況(2022 年度).2023
(2)農林水産省.野生鳥獣被害防止マニュアル -総合対策編-.2023
(3)堂山宗一郎.ニホンジカはどのようにして柵内に侵入するのか?.
JATAFFジャーナル7.2019
(4)長門勇治,吉仲怜.鳥獣被害対策における電気柵管理の実態と方向性.
農業経営研究49.2011
(5)村松広貴ら.防護柵の延伸がイノシシによる水稲被害軽減に直結しない理由.
野生生物と社会7.2019
(6)山端直人ら.獣害対策の継続が集落のソーシャル・キャピタルに及ぼす影響.
農村計画学会誌34.2015
(7)岡崎(田中)麻衣子ら.クズ(Puerarialobata(Willd.) Ohwi)登攀茎の生長と莢生産.
日緑工誌43.2018
(8)高橋満彦.鳥獣法の根本は変わるのか…2014年法改正を題材に.
ワイルドライフフォーラム春夏号.2014
(9)松尾浩司(環境省).狩猟免許を有しない従事者制度の概要.
地域ぐるみで取り組む鳥獣被害対策に関する研修会発表資料.2013
(10)辻井修ら.階層分析法による果樹の鳥獣対策に関する感性的評価.
愛知農総試研報53.2021
(11)農研機構・鳥獣害管理プロジェクト.
果樹園のカラス対策「くぐれんテグス君」設置マニュアル.2013
(12)埼玉県農業技術研究センター.楽落ライト設置マニュアル.2018
(13)環境省.鳥獣の保護を図るための事業を実施するための基本的な指針
(平成28年10月告知版).2016
(14)小坂井千夏.廃果場,エネルギー獲得効率の高い鳥獣の餌場
-鳥獣害防止に向けて必要なこと-.JATAFFジャーナル7.2019
(15)Sophie Rossi et al. Controlling of CSFV in European wild boar
using oral vaccination: a review. Frontiers in Microbiology6. 2015
土のはなし-第31回
危機に瀕する世界の土-その1
古代文明の崩壊と土の劣化
前 ジェイカムアグリ株式会社
北海道支店 技術顧問
松中 照夫
連載の第25回から前回まで,農業はそのやり方次第で環境に悪影響を与えることがあると述べてきた。したがって,農業を環境保全的に営むことが重要となる。農業は人の食料を生産するがゆえに,止めるわけにいかないからである。
農業の作物生産の場を支えているのは土である。しかし,その土が荒廃して作物生産に適さなくなってしまう現象(これを土の劣化という)が地球規模で広まり,すでに土の3分の1が劣化しているという(Silva,2014)。まさに土が危機に瀕している。それはなぜなのか,土の劣化の現状や原因を今回からしばらく探ってみたい。
1.土への無関心は文明を崩壊させる
およそ1万年前,原始人は農業によって食料を確保する知恵を得た。農業で食料の安定生産の基盤ができると,人口扶養能力が高まった。しかし皮肉なことに,それによって狩猟採集の生活に後戻りできなくなった。増えた人口の食料を狩猟採集では支えきれないからである。農耕定住生活は,自然生態系からはなれて人工の生態系をつくり,集落をつくってしだいに繁栄していった。ここに古代文明の芽ができた。
しかしこの文明の繁栄は,30から70世代(800 から2000年)以上長続きすることはなかった。その根本原因は,自然生態系を無視し,土の肥沃度維持に無関心だったため,食料を持続的に生産できなくなったことにあると,カーターとデール (1975)はその名著「土と文明」で指摘している。
彼らの指摘の一部を以下で見てみよう。
1)肥沃な三日月地帯,メソポタミアの例
「肥沃な三日月地帯」とは,チグリス川とユーフラテス川の流域に,その上流地帯からヨルダン川流域,死海までの低地帯のことをいう。かつてこの地域は「乳と蜜の流れる土地」とさえいわれた肥沃な土があった。この地域は降水量が少ない。
そのため,作物栽培にはチグリス,ユーフラテス両河川から水を引き込むかんがい(灌漑)が必須だった。両河川上流にあるアルメニア高原は豊かな森林だった。しかし文明に触発されて人が集まると,燃料や建材用に森林が伐採され,家畜が過放牧された。森林を失った高原の土は水を保持しきれず,表土が侵食されて土砂が河川に流れ込み,かんがい用水路に堆積した。用水路の機能を維持するのは,奴隷労働による土砂の除去だった。
しかし,メソポタミアを攻撃する民族は,かんがい用水路に無関心だった。やがて用水路は土砂でふさがれ,使用不能になる。決定的な打撃は,遊牧民の蒙古人がこの地域を襲った時だった。遊牧民はかんがいの重要性が理解できず,用水路を完全に破壊した。さらによくないことに,蒸発の盛んなこの地域では,かんがいによって地下水と地表がつながると,おびただしい塩類が地表に導かれ蓄積し,土が塩類化した。塩類化は土の作物生産力を完全に失わせた。
食料生産に不可欠なかんがい水確保の水路が土砂で埋められたこと,土が塩類化しやすかったこと,これらの要因がメソポタミアでの食料の持続的安定供給を困難にして,人口扶養能力を低下させた。こうして文明が衰微していった。

2)エジプト・ナイル川流域の例
自然を生かした土の肥沃度の維持:同じ古代文明発祥の地でもエジプトのナイル川流域は,メソポタミアとは事情がちがう。ナイル川は毎年正確な周期で増水と減水をくり返す。おもな水源が,エチオピア高原と中央アフリカの高地の雪解け水だったからである。年に一度,夏に必ずやってくる洪水は,古代エジプトに実りをもたらした。
まず,ナイル川の流域に沿って,水田の畦畔のような囲みをつくっておく。そこへ,増水期で溢れ出た泥水が田植え前の水田のように蓄えられる(湛水かんがいという)。数週間その状態を維持すると,泥水に含まれる肥沃なシルト(微砂)分が地面に沈殿し,土にも水が十分に浸透する。その後,余剰水を排水し,肥沃な泥土にコムギなどの種子を播く。自然との調和を考えた当時の人々は,増水期のナイルの巨大な水のエネルギーを征服しようとしなかった。むしろ自然を上手に利用した湛水かんがいの技術を確立した。しかし,この技術にも欠点があった。それは,年1回のナイルの増水期しか作付けできない弱みだった。
通年かんがいへの転機:19世紀以降になると,エジプトでは綿花の輸出が計画された。しかし,綿花はナイル川の減水期(3~4月)に播種され, 10月に収穫される夏作物である。このため,増水期の水を利用できない。そこで,減水期の低い水位の水を利用するため,深いかんがい用運河が掘られた。こうして,ナイルの水量の増減に関係なく,水を通年利用する環境が整った。
この通年かんがいへの熱い思いが,後の1903年,イギリスの援助によるアスワンダムを完成させた。このダムの利用で安定した農業生産が約束された。そのおかげで人口が大きく増え,1882年に 700万人だった人口が,70年後の1952年には2,000 万人に達した。ところが,人の生活のための土地利用が進むと,原流域の山林が伐採され,家畜の過放牧もはじまった。こうした土地利用の変化で,ダムに運ばれる土砂が多くなり,ダムは土砂で徐々に埋められ,貯水機能が低下した。 ダムの機能低下はかんがい不能をもたらし,作物生産の減少につながった。そこでアスワン・ハイダムの建設が,ソビエト(現在のロシア)の援助で1960年に着工,10年後に完成した。これによって通年かんがいの農地ができ,作付けも年に数回可能となり,作付面積が大幅に拡大した。
ダムの功罪と自然の摂理:アスワン・ハイダムの完成は水問題を解決したかにみえた。しかし,張り巡らされたかんがい用水路に水生カタツムリが大発生,それを中間宿主とする寄生虫ビルハルツ住血吸虫の蔓延と,土の塩類化が始まった。
年に一度のナイルの洪水は,地表に蓄積されがちな塩類を洗い流してくれた。しかも,養分に富む肥沃なシルトを上流から運び,古い土の上に堆積してくれる。これによって,土の塩類化を自然に克服し,土の肥沃度が維持された。これが「肥沃なナイルの低地土」を裏付ける古くからの自然の摂理であった。
しかし近代になって,人々はナイルの流れを自分たちで制御しようと試みた。農業安定化のために築かれたはずのアスワン・ハイダムは塩害と寄生虫をもたらした。自然の摂理を生かしきれなかったかんがい農業の悲劇である。
2.教訓をどう生かすか
古代文明の衰退が教えることは,食料生産の基盤である土を保全しないで食料の確保はあり得ず,高度な文明も維持できないという事実である。太平洋の孤島イースター島での資源の枯渇による文明の崩壊もまた,現代の私たちに貴重な教訓を提供している(ポンティング,1994)。これらの教訓をどう生かすか,現代の私たちに問われている。
