
第750号 2023(R05).05発行
PDF版はこちら
§メロンとキュウリのマグネシウム,カリウムの欠乏症状と再移動
元 岡山大学大学院 自然科学研究科
桝田 正治
§土のはなし-第21回
「土は生きている」といわれるのはなぜ?
-土は生き物なのか
前 ジェイカムアグリ株式会社
北海道支店 技術顧問
松中 照夫
メロンとキュウリのマグネシウム,カリウムの
欠乏症状と再移動
元 岡山大学大学院 自然科学研究科
桝田 正治
はじめに
野菜の生理障害について,筆者は発生部位と生育段階ならびにその発生要因を一つの表にまとめて示した7)。それから約20年経った現在においても,それらの症状の要因に新しい知見は加えられていない。
このことは近年,野菜の生理障害があまり大きな問題となるには至っていないことを示唆する。種子繁殖性の野菜では,新品種の生育特性や連作に伴う病害の研究が多く生理学的研究そのものが少なくなったように感じる。栄養繁殖性のサツマイモ,サトイモ,ジャガイモ,ニンニク,イチゴなどはウイルスフリー原種を保存し,それから増殖させ種苗として市販されるが,通常,栽培者が1,2年繁殖していると新しくウイルスに感染してしまい,外見上の症状からは,病的あるいは生理的としてその要因を見分けることが難しくなる。勿論,マグネシウム,カルシウムの欠乏や拮抗阻害ならびに微量要素の欠乏によるものも少なくない。
Mg欠乏症は微量の亜硝酸ガスやアンモニアガスによって引き起こされる障害にも時には類似すること,さらには光化学オキシダントやハダニの害と酷似するものも多く,その要因をすぐには判断しにくい場合が多い。
前者のガス障害はハウス栽培において,特に,油かすや鶏ふん等の有機肥料の多用で発生しやすい。また,ホウレンソウ,大根,小松菜など直播する野菜では葉が地表に接するので露地での栽培でも障害は現れやすい。ガス発生のメカニズムはすでに明らかにされており,ガス化を抑えるためには土壌pHの酸度調整ならびに窒素肥料,とりわけアンモニア態窒素肥料の施与量に注意する必要があるとされる。
後者については,図1の①~③に示したように,光化学オキシダント障害,強光や高温障害などがあげられる。これらは一般に知られる写真④のメロンの葉脈間クロロシスとも類似していることがある。このメロン着果節葉に発生する症状とMg欠乏との関連性を本稿では論じようとするものである。光化学オキシダントは工場や自動車などから大気中に排出される窒素酸化物(NOx-ノックスと読む)や揮発性有機化合物が紫外線を受けて生成される物質(オゾン(O3)やパーオキシ・アセチル・ナイトレート(PAN)など)が動植物の細胞に障害をもたらす。

図1の写真①に示したアサガオは,その障害の典型例で1970年代の光化学スモッグ,50年ほど前に筆者も京大付属農場で調査したが,当時これは全国的に取り上げられた指標植物である。この時に,野菜ではサトイモが敏感に反応することを知った。最近でも,キュウリの葉が初夏の強光に一時的に遭遇すると翌々日に葉脈間が斑点状に白化する現象が見られる。これは恐らく過剰な光エネルギーを活性酸素消去系が瞬時に処理しきれないために生じる現象であろうと推測されるが,エビデンスは得られていない。
写真②に示したアールスメロンでも高温と一時的な強光により葉脈間の一部が一夜にして変色する。アールスメロンは高温耐性の非常に高い作物であるが(43℃になっても強光の反対側にある葉では障害は生じない),高温に強光が加わると葉に障害が発生することになる。
写真③に示したダニ害は,亜硝酸やアンモニアガスおよび光化学オキシダントや強光による障害とは異なり,葉にかすれ症状が生じるのが特徴的で,進行すると葉の裏に肉眼でも多くのダニを見ることができる。しかし,初期の症状はMg欠乏症と類似しており,なかなか識別しにくい。特に,高温期にさしかかる頃の野菜では生理障害よりもまずダニ害を疑ってみるとよい。
メロンの葉枯れ症およびキュウリのMg欠乏症状と成分移動
1)メロン葉枯れ症の概要
一般にウリ科作物では生育中期以降,特に果実の肥大が進む頃になると近傍の葉の葉脈間にクロロシスを発現することが多い。1980年代には各地でプリンスメロンの葉枯れ症が多発し多くの試験場での研究報告はMg欠乏によるものとした。そんな中で兵庫県農業総合センターの津高12)は葉枯れ症がK欠乏によって生じていると報じた。筆者の知る限りではK欠乏とした論文は後にも先にもこれが唯一である。
藤本2)はハウスメロンの葉位別Mg含有率を正常株と葉枯れ症株について調査し,図2の結果を得ている。果実は第13節~16節に発生した側枝に着果させ,下位葉から上位葉まで主茎節位の葉分析を行ったもので,13節~16節のMg含有率は正常株では0.3%以上,発生株では0.2%以下となっていることが分かる。この結果から,同氏は葉枯れ症の発現レベルは,葉中Mg含有率で0.2%レベルにあるとし,葉枯れ症は果実の肥大成熟に伴って葉のマグネシウムが果実へ転流し,その結果,葉の葉脈間が黄化し,やがて黄褐色を呈して枯れるが,このタイプのものは成熟の目安とされることもあるほどで収量,品質にほとんど影響しないと述べている。
筆者4)も自根キュウリの生理障害,褐色小班葉が0.19%以下で発生すること,欠乏症状を呈した葉のMg含有率は正常葉のそれに比べて確かに低いことを認めている。しかし,これらの結果は,葉から果実へマグネシウムが移行することを示すものではない。従来,マグネシウムは移動しやすい成分とされているが3,10,13),本当に移動しやすい成分なのであろうか?
2)キュウリ樹体内における成分移動の難易性
そこでまず,筆者は養分除去に伴う水耕キュウリの成分変化,とくにマグネシウムについて他の成分と比較した。園試標準培養液で育つキュウリ〈主枝22節,側枝3節で摘心〉の培養液を水道水へ入れ替え,第5節の葉と根の乾物重並びに含有率を経時的に調査した。乾物含有率からみた根の減少率は窒素とリンが大きく,水処理4日後で61%,カリウムとマグネシウムは,さらに減少し続け処理8日後には減少率74%,82%となった。
一方,葉においても窒素,リン,カリウムは大きく減少した。特に,リンとカリウムの減少率が著しく,それぞれ処理8日後には当初の60%,63%となった。これに対して,カルシウムとマグネシウムは,わずかに減少するに過ぎなかった5)。勿論,処理期間中に光合成により乾物重は増加する。その時の乾物率から葉における含有量を求め処理開始時を基準に相対比として示したのが図3である。
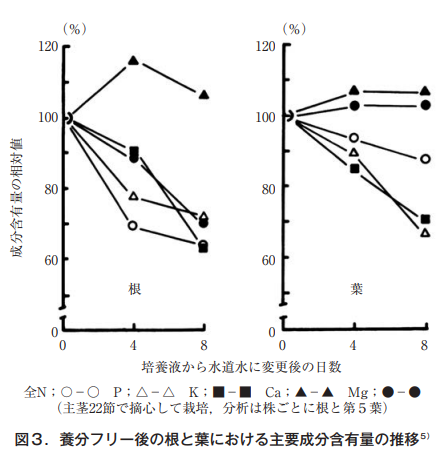
根についてみると,カルシウムは全く変化せず,減少率は全N=K>P=Mg>Caとなった。一方,葉ではカルシウム,マグネシウムは全く変化せず,減少率はP=K>全N>Mg=Caとなった。よく移動するとされるマグネシウムに変化が見られないことは,溶液の養分除去(水道水)が8日間と短かったことを考慮しても,キュウリ葉中のマグネシウムは,少なくとも窒素,リン,カリウムに比較して移動しにくい要素であるといえる。
3)キュウリ個葉に散布したマグネシウムは,その葉でも移動しない
次に,生育中のキュウリを摘心し2本仕立てとし,草丈約70㎝に達した段階で培養液からマグネシウムを除き,それから2日毎に4回,2本仕立ての一方に1%MgSO4,Mg(NO3)2,あるいはMgCl2の溶液を樹全体に散布したところ,いずれの処理でも葉脈間クロロシスは完全に抑制できた(対照区:無散布の樹では発現する)。この散布効果が葉の表裏で変わらないことから,散布Mgはクチクラから細胞内に入り,そこで種々の形態に変化するものと考えられる。
そこで,再度,欠乏症が発現すると予想される中位葉の1枚に散布処理を試みたところ,散布葉そのものは緑色を保ったが,その上位節と下位節の葉はクロロシスを呈した(写真省略)。このことは散布したマグネシウムがいかに動きにくいかを示すものである。
そこで更に,個葉におけるMg移動の難易を知るため,クロロシスを発症すると予想される中位葉1枚を特定し,底を開けた紙コップを葉の一部に押し当てその中に1%MgSO4溶液を1日おきに4回散布した。その結果を図4に示す。
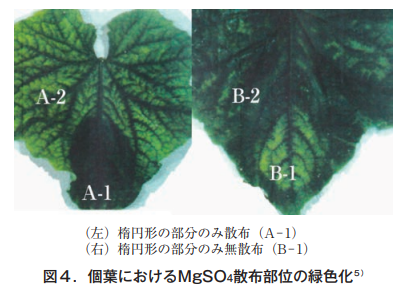
散布部分は緑色が保たれるが,無散布部位は典型的な葉脈間クロロシスを呈する。これらの部位におけるMg含有率を表1に示した。
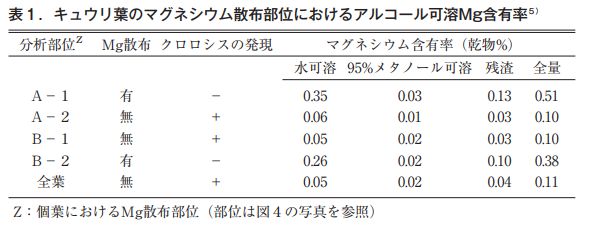
散布部位A-1,B-2の水溶性Mgは,無散布部位A-2,B-1の約6倍で散布Mgが葉に浸透していることは明らかで,無散布部位の含有率が対照として設けた全葉無散布の含有率と同じ値を示すことから,少なくとも散布Mgは個葉において移動しないといえる。
また,散布部位と無散布部位で葉色に差が認められるにも関わらず,クロロフィルの骨格にあるマグネシウム, すなわち95%メタノール可溶性Mgには差が認められない。このことは,クロロフィルを構成するマグネシウムは通常の機器では測定不能なほど微量であることを示唆している。
4)キュウリの白変葉とグリーンリング葉はアンモニア態窒素によって誘発される
接ぎ木キュウリ〈台木はカボチャ〉の白変葉が,1970年代後半から高知,宮崎で多発し十数年にわたり多くの研究者がその原因の究明にかかわった。当初,この症状はMg欠乏によるとする報告が多く発表されたが,これは,いわゆる黄化(クロロシス)ではなく白色壊死(ネクロシス)することから,Mg欠乏だけでは説明できない現象であった。多くは葉縁が緑色に保たれていることから’グリーンリング葉’と称され,激しい時は一夜のうちに褐変,光が当たると白変が形成される。このような葉の発生要因を,高知県園芸試験場の松本ら8)は,見事に解明したのである。簡単に要約すれば以下のようになる。
当時のハウス栽培では土壌消毒剤として臭化メチルが使用されていた。この土壌に有機肥料の油かすが施与される。油かすは微生物(アンモニア化成菌)の働きでアンモニア態窒素となる。アンモニア態窒素は硝酸化成菌によって硝酸態窒素に変わるが,低地温下では,硝酸化成菌の活性が低いため(硝酸化成菌はアンモニア化成菌より低温に弱いことは一般に知られている)アンモニア態窒素の濃度が高まる。さらに,臭化メチルが処理された土壌では硝酸化成菌の活性がより弱まりアンモニア態窒素の集積が顕著となる。土壌中にマグネシウムが存在してもアンモニア態窒素が高いと拮抗作用で吸収が抑制される。キュウリの葉のグリーンリング葉は土壌のアンモニア態窒素の過剰とマグネシウムの吸収阻害によって引き起こされるとしたのである。
それでもなお筆者は,キュウリの白変葉の誘発要因を証明するには,実際に農家のハウスで見た症状を自ら再現する必要があるとの思いから,自根キュウリと接ぎ木キュウリの水耕栽培において試行錯誤を重ねた。その結果,養液のアンモニア態窒素の濃度を高め,マグネシウムの濃度を低めた時にカボチャ台木のキュウリに白変葉およびグリーンリング葉が発現することを認めた(図5)。

なお,専門用語として白変葉(interveinal necroticleaves)は改定園芸学会用語集〈2004〉で初めて取り上げられ,グリーンリング葉は白変葉の一部とした。
5)メロンの着果に伴う葉の障害とMg・K含有率の変動-マグネシウムが再移動するのは確かであろうか?
一般に,メロンは収穫期に近づくと果実近傍の葉にクロロシスが生じる。これが収穫期の目安として古くから語られてきた。教科書的にはこの症状は葉から果実へマグネシウムが移行するためであると記載されている。筆者も,大学の講義でそのように習ったような気がしている。アールスフェボリットでは,着果後40日ころから葉脈間がわずかに黄化しはじめ,50日頃には激しくなり褐変し始めることが多い。はたしてこれは,葉から果実へマグネシウムが移行することによって発現する症状であろうか?
そこで,果実の有無と葉の障害程度を調べるため,果実を着果後40,50,60日で摘果し70日後に各株から果実節葉を採取して分析した6)。メロンの着果枝の基本型は,図6の右に示したように,主茎第13節から伸びた側枝〈子蔓〉の第1節に雌花が着く。近傍の葉をA,B,Cとして左にその形状と色を示す。上段は,着果後40日目に果実を除去した株(30日間は果実無し),下段は最後まで(70日間)果実を残した株,それぞれの葉色である。
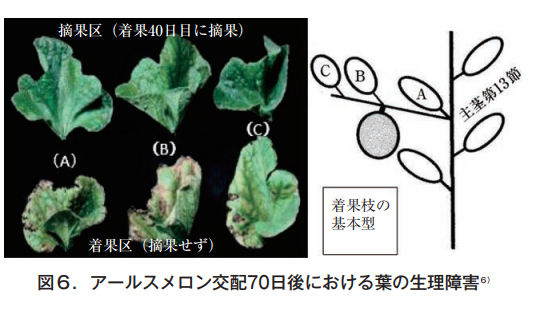
写真からも明らかなように40日目に摘果した場合は,いずれの葉も緑色を保つが,摘果しないと葉は変形し,葉脈間はクロロシス,葉縁はネクロシスを起こす。その症状は着果節葉で著しい。通常,果実は55日前後で収穫されるため,ここまで厳しくはならないが,この試験では果実の有無との関係を明らかにするため着果期間を70日間と長く設定した。これらの葉のマグネシウム,カルシウムおよびカリウムの含有率を表2に示した。
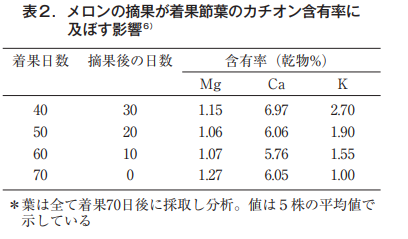
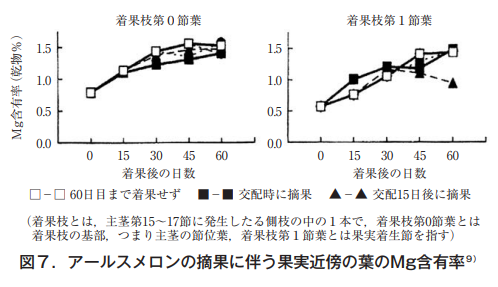
マグネシウムは果実の有無にかかわらず,その値は1.1%~1.2%台でほとんど変化していない。カルシウムは若干低下する傾向にある。これに対して,カリウムは果実が長く存在するほど葉での含有率は低下し,70日間果実を残した株は40日目で除いた株の1/2以下となった。西村ら9)も着果時から経時的に葉分析を行いMg含有率は低下するどころか,むしろ徐々に高まることを明らかにした(図7)。
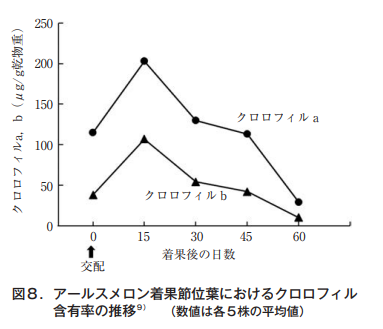
通常,葉は着果40日頃から黄色を帯び始めるが,この変化はクロロフィル値でも知ることができる(図8)。交配15日目に葉色は最も濃くなり,その後,急速に薄れていくことが伺える。
筆者が岡山大学に赴任する前の蔬菜園芸学研究室(益田忠雄教授)では,研究テーマの多くがメロンに設定されていたようである。当時5年間の卒論からマグネシウム,カルシウムおよびカリウムのデータを取り出し成分別に画いてみた。それが図9である。
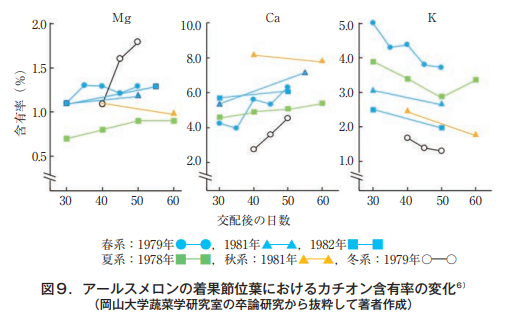
これからも分かるように,マグネシウムとカルシウムは果実肥大につれて徐々に上昇する傾向にあるが,カリウムは明らかに減少する。交配後50~60日には,当時も葉脈間クロロシスや葉縁ネクロシスが発現していたであろうが,葉に現れる現象と分析値の関係については考察されていない。着果近傍の葉を観察していると,メロンのMg欠乏による葉脈間クロロシスは収穫期に近づくにつれて徐々に激しくなることから,葉のマグネシウムが果実に移行するとする見解は自然の理にかなっており,何ら不思議ではない。
しかし,今もって果実に移行するとするエビデンスは得られていない。現下のルーラル電子図書館の農業技術事典(農研機構)にもマグネシウムは移動しやすい成分で果実の肥大・成熟にともなって葉から果実へ移動し,果実周辺の葉に欠乏症状が現われることがあると記載されている。
おわりに
野菜の栄養生理,とりわけ養分欠乏症は完全培養液から一つの成分を除いたときに同症状が出るかどうか,除かなくても他の成分を高めたときに同症状が出るかどうか,この2つの条件で研究されることが多い。図10に示したように,根における養分吸収にはイオンが相互に影響しあうため葉の欠乏症状を成分一つに帰すのは難しい。
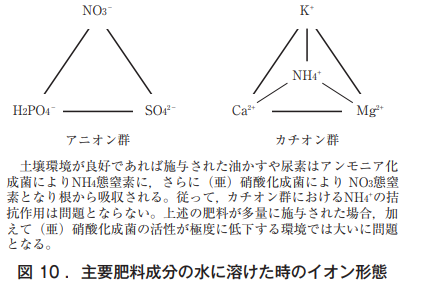
植物の根は,アニオン群,カチオン群から養分を選択的に吸収するが,木部液でのイオン総量〈meq〉は,プラスイオンとマイナスイオンで釣り合っている。土中にはNa⁺,Cl⁻,HCO3⁻などを含め多種の微量要素イオンも存在する。なお,カチオンでは一価が二価よりも吸収されやすいとされ,それらの拮抗作用も知られている(NH4⁺は筆者の栽培経験から挿入)。キュウリの白変葉はカボチャ台木で,かつNH4-N過剰下のMg欠乏により生じ,Mg濃度 が少し高いとグリーンリング葉になりやすい。つまり,白変葉は二つの要素により引き起こされる生理障害であり,一夜にして壊死症状を呈することから,筆者はアンモニアが直接関与している可能性が高いと推察している。
なお,同様な現象はトマトの尻ぐされ症にも認められる。トマトの尻ぐされ症がCa欠乏によって生じることは既知の事実となっているが,NH4-N過剰下のCa欠乏では激しい壊疽(えそ)が果頂部に現れる。2因子以上の要因による生理障害,メロンの着果節の障害葉もクロロフィルの崩壊とカリウムの再移動によって引き起こされる可能性が高いと推察される。
いずれの成分においても果実への再移動に関しては放射性トレーサーを用いてそのエビデンスを得る必要がある。Bukovacら1)は,葉面散布した28Mgは45Caと同様に移動しにくいと記している(ただし,放射性Mgについてはデータ無し)。本稿のキュウリ個葉の散布実験と分析値からマグネシウムは隣の細胞にすら移動しにくいことが確認できた。
最近,田野井11)は,総説「植物のミネラル輸送研究最前線」において,Mg2+トランスポーター遺伝子(MGT)のファミリーがシロイヌナズナ,イネでそれぞれ9個,トウモロコシで12個報告されているとし,それらのマグネシウムの輸送機構について解説しているが,メロンの葉におけるMGT発現量が散布したマグネシウムによって増大しなければ,もしくは,もともと葉におけるMGT機能が微弱であればマグネシウムはその部位に止まり移動しないのも当然と言える。メロン果実近傍の葉の葉脈間クロロシスはクロロフィルの崩壊によって生じているが,葉緑体でフリーとなったマグネシウムが果実に移動しないのもMGT遺伝子の発現量が関係しているかも知れない。
一般に,成熟葉ではマグネシウムの約半分が葉緑体に存在するとされ,その一部が葉緑素構成のマグネシウムであることを考えると,再移動に関してはそのわずかな量を問題にしなければならない。とはいえ,クロロフィル崩壊により生じたフリーのマグネシウムが果実へ再移動するとするには,これまでの実験結果からどう考えても無理がある。28Mgの半減期が約20時間と短いため実験上の制約を受けるにしてもトレーサー法は動態解析に欠かせない手法と言えよう。
最後に,もう一点不思議に思うことは,収穫期の近づいた果実近傍の葉になぜクロロシスが生じるのであろうか?果実が存在しなければ,それは生じない。この頃には果実から何らかのシグナルが葉に送られているはずである。それはクロロフィル崩壊のシグナルといえるが,このシグナルはメロンにとって何を意味しているのであろうか?Whole plant physiology の観点からも今後の研究に期待したい。
引 用 文 献
1.Bukovac,M.J. and S.H.Wittwer. 1957.
Absorption and mobility of foliar applied nutrients.
Plant Physio. 32:428-435.
2.藤本順子.2008.
園芸作物における栄養障害診断手法の開発と防止対策に関する研究.
島根農技研報. 8:1-45.
3.Jacob,A. 1958.
Magnesium: The fifth majorplant nutrient. 34-54.
Staples Printers Limited,Kingdom.
4.桝田正治.1984.
接ぎ木・自根キュウリの高K培地と高NH4-N培地で発生するMg欠乏症の差異.
農業及園芸59:1051-1053.
5.桝田正治.1989.
キュウリ葉におけるマグネシウムの欠乏症と移動の難易.
岡山大学農学報74:7-14.
6.桝田正治・金沢孝美.1993.
メロン着果節葉のカチオン含有率と養分欠乏症.
園芸学会中四支部大会要旨集32:31p.
7.桝田正治・寺林 敏.2003.
生産技術. 図説野菜新書.矢澤 進(編).97-151.
朝倉書店.東京.〈131pに記載〉.
8.松本満夫・上杉郁夫・柳井利夫:1981.
施設栽培における接ぎ木キュウリのMg欠乏症(グリーンリング症).I.MB剤による硝化抑制が養分吸収とグリーンリング症に及ぼす影響.
高知農林報.13:1-10.
9.西村安代・福元康文・島崎一彦.2004.
アーメロン(Cucumis melo L.)の葉内無機成分に及ぼす着果の影響.
生物環境調節,42:137-146.
10.嶋田典司.1976.
マグネシウムの生理作用.土壌肥料作物栄養大辞典(第3版).102-105.
養賢堂.東京.
11.田野井慶太朗.2021.
マグネシウムの輸送機構.
日本土壌肥料学雑誌.92:108-113.
12.津高寿和.1982.
プリンスメロンの葉枯れ症の原因と対策.
農業及園芸57:1162-1166.
13.山崎 伝.1975.
微量要素と多量要素〈第7版〉.175-183.
博友社.東京.
土のはなし-第21回
「土は生きている」といわれるのはなぜ?
-土は生き物なのか
前 ジェイカムアグリ株式会社
北海道支店 技術顧問
松中 照夫
今回の話題はこれまでとがらりとかわり,土そのものに目を向けてみる。農業を安定して支えているのは土である。その土は,私たちの命を守る食べものを生産する場であると同時に,さまざまな生き物の命も育んでいる。こうしたことから,土への畏敬の念をこめ,しばしば「土は生きている」といわれる。それはどうしてなのだろう。
1.土は生き物なのか?
土が本当に「生きている」,すなわち生き物なのかと改めて問いただされると,土が生き物であるという人は多くはないだろう。
一般的な概念で生き物というのは,多細胞生物である。その多細胞生物であるためには,①分化と生長,②繁殖と遺伝,③環境変化への自律性を満たす必要がある(岡島,1989)。しかし,土に両親がいて,その遺伝的要素を引き継いで大きくなり,子供を育て,死んでいくとはだれも考えないだろう。「土は生きている」というのは,あくまでも,土を譬喩(ひゆ)的に表現しているにすぎない。土を敬愛するあまりに,土が生き物であると混同するのはつつしむべきだろう。
しかし,それが譬喩であったとしても「土は生きている」という表現は,私たちの心を捕らえてはなさない。それはなぜなのか。
2.生き物の自律性に似た性質が土にもある
土には,生き物が持つ環境に対する自律性,すなわち,外部から刺激を受けても,自分自身で状態を維持しようとする働きを感じさせる性質がある。それが土の緩衝力である。
具体的に土の緩衝力を見てみよう。土に酸性物質として塩酸,アルカリ性物質として水酸化ナトリウムをそれぞれ滴下する。これによって,土は外部から刺激が加えられたことになる。この時,図1に示したように,純水(H₂O)の場合,ほんのわずかな滴下量で,pHが酸性側にもアルカリ性側にも大きく変化する。
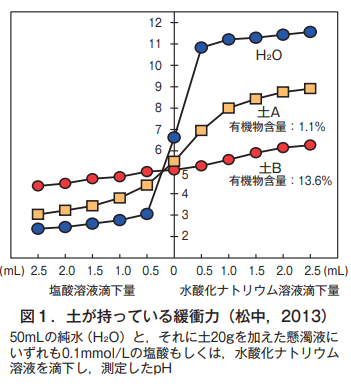
一方,土Aと土Bのどちらも,純水より変化の幅が小さい。つまり,土は純水より緩衝力が大きいのがわかる。ただし,有機物の少ない土Aは,有機物の多い土Bよりも変化の幅が大きく,したがって緩衝力は小さい。土の有機物は,酸性の原因である水素イオン(H⁺)やアルカリ性の原因である水酸イオン(OH⁻)を静電気的に保持する能力を持つ(土の静電気的なイオン保持能については,昨年の2月号の第9回を参照)。このため,有機物がそれらのイオンを保持して動きを抑え,外界からの刺激をやわらげることができる。土Bのほうが土Aよりも緩衝力が大きいのは,土の有機物含量の違いに基づく。このような土が持つ緩衝力は,生き物の自律性とよく似ている。
しかし,土の緩衝力だけが「土は生きている」といわれる要因ではないように思う。土で生活する生き物たちの働きは,私たちの目に見えない現象であることが多く,あたかも土自身がおこなった活動のように見える。これも「土は生きている」と感じさせる大きな要因だろう。
3.生き物たちの生活の証し-土壌呼吸
土に与えられた堆肥が作物の栄養分となること,また,秋になって土の上に落ちた木の葉や,台所から出た生ゴミを土に埋めておくとそのうち姿を消すことなどは,いずれも,それら有機物を土の生き物たちが分解した結果である。
土の生き物たちが,堆肥や落ち葉,生ゴミなどの有機物を分解すると,分解産物の一つとして二酸化炭素(CO₂)が排出される。この様子は動物の呼吸に似ていることから,「土壌呼吸」といわれる。あたかも土が呼吸しているかのような感覚である。しかしこれは,あくまでも土の中で生活している生き物たちの活動の証しであって,土自身が呼吸しているわけではない。
4.有機物分解は土の生き物たちの連係プレー
上で述べた落ち葉を例に,土の生き物たちが有機物を分解する時の,みごとな連係プレーを見てみよう(図2)。
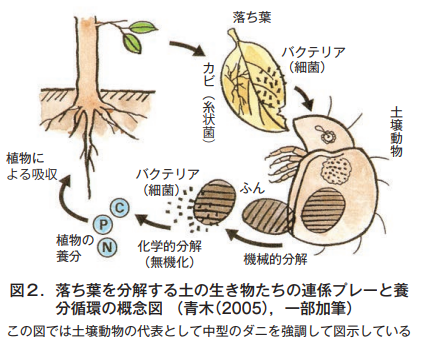
土の表面にある落ち葉(植物遺体)は,そのまま乾いた状態では大きく変化しない。しかし,それがひとたび雨に打たれて濡れると,そこにまずバクテリア(細菌)やカビ(糸状菌)がとりつき,この植物遺体をある程度軟らかくする。
すると今度は,ミミズやワラジムシ,ダンゴムシなどの大型の土壌動物が登場し,植物遺体をエサとして食べて粉砕するとともに,植物遺体や食べかすを土の中に引きずり込む。その後はダニやトビムシなど中型の土壌動物が担当し,引きずり込まれた有機物をエサとして食べ栄養分を体に取り込み,不要なものをふんとして排泄する。
この排泄されたふんや,引きずり込まれた植物遺体もバクテリアやカビがエサとして食べ,最終的に二酸化炭素や水,そして無機物に変化させる。これによってできた無機物は,再びそこで育つ植物の栄養分として根から吸収される。こうして養分の循環が成立する。もちろん,動物の遺体についても植物とまったく同じしくみである。
土の中でおこるさまざまな物質変化には,土の生き物たちが関わる変化が含まれている。したがって土の中での物質の変化を語るとき,土の働きと土の中の生き物の働きをあいまいにして述べると,両者が混同されてしまう。
5.土の働きと生き物の働きを区別する
土の働きと表現しても,それは土に生き物のような「意志=合目的性」があって,土自身の意志が能動的に働くことを意味していない。「土が作物に養分を供給する」という表現も実は不適切で,土自体が作物へ意図的に養分を供給することはない。土の中の水(土壌溶液)に溶けているさまざまな養分イオンの中から,作物が必要とする養分,例えば陽イオンの養分を選択して吸収すると,それまでの電気的中性原理(陽イオンと陰イオンの荷電量が等しい状態)のバランスがくずれる。くずれたバランスをもとの状態に戻すように,土の中で多様な反作用がくり広げられ,電気的中性原理を維持するために陽イオンが土壌溶液に放出される。
これが「土が作物に養分を供給する」と表現される現象である。この反作用も一定の原理(この例では溶液での電気的中性原理)によって律せられており,生き物がもつ自由な意志がはいり込む余地はない。
土と土の中の生き物たちの働きを区別した上で,「土は生きている」という譬喩の真意を理解したいと思う。